מדוע כדאי לך לדעת על עבודתו של הרולד הילמן על התא החי?
מאת: מיה בריז (Mia Breeze) | פורסם באנגלית בפברואר 2024
תרגום לעברית ועריכה: גדעון כינר. הערות ב- [סוגריים מרובעים] הן מאת העורך | פורסם בעברית: יולי 2024
מקור: https://dpl003.substack.com/p/why-you-should-know-about-harold
מאמר זה בוחן כיצד ביקורתו של הרולד הילמן על [מחקר] התא החי בביולוגיה, משפיעה על ההבנה הנוכחית שלנו את מה שנקרא "חומצה דֶה-אוֹקסִי-רִיבוֹ-נוּקלֵאית" (דנ"א), ואת הנושאים של גנטיקה, וירוסים, אורגניזמים מהונדסים גנטית (GMO) והבנתנו את היצורים החיים.
ניסיתי לתמצת ולפשט ככל האפשר את עבודתו של הילמן הנוגעת לתא החי כדי להפוך אותה לנגישה יותר לקורא הממוצע, אך בהתחשב בנטייתו של המדע המודרני לסבך ולטשטש את הדברים, יש כאן הרבה קשרים להתיר.
לכן, אם אתם מסוגלים לשים בצד את טווח הקשב המודרני המקוצר, ולהשקיע זמן בקריאת מאמר זה, סביר להניח כי לכשתסיימו לקוראו, תדעו בביטחון מה ניתן באמת לדעת על נושאים אלה ומה כנראה שגוי.

הערה לגבי תרגום ועריכה:
קצת מסובך לבחור כמה להתערב במבנה מאמר במהלך תרגומו. מצד אחד אני רוצה להישאר נאמן למה שהתכוונה הכותבת לומר, ומצד שני להגיע לבהירות מירבית של הנאמר, בצורה כזו שיהיה מובן וברור לקוראיו בעברית. אני מקווה שהבחירות שלי כאן מצליחות להגיע לאיזון ראוי בין השיקולים הללו. מוזמנים לכתוב לי ולהעיר דרך צרו איתנו קשר אם מצאתם שלא כך הדבר (בציטוט הנוסח השגוי, לדעתכם), ואתקן בהקדם את מה שאכן נדרש לו תיקון.
כל הכתוב בתוך [סוגריים מרובעים] התווסף בעריכה לטובת בהירות הכתוב.
תוכן עניינים
1. הרולד הילמן
בהספד על הרולד הילמן (לונדון 1930 – 2016) אנו למדים מאחיו כי:
"… הרולד הילמן, … היה מדען ביולוג שמחקריו … אתגרו שוב ושוב את הקהילה המדעית האורתודוקסית בפרשנויותיה להשפעות של מיצוי, ייבוש וצביעה [של דגימות] על מבנה התא [בעת הכנת דגימה] לצפייה במיקרוסקופ אלקטרונים.
בספר "התא החי" (1980), שכתב הרולד יחד עם פיטר סארטורי, הוא טען שתהליך ההכנה גרם לכשעצמו לשינויים משמעותיים בתאים עצמם, ולכן אינו אמין, דעה זו נדחתה על ידי רוב עמיתיו, ועליה שילם מחיר יקר במונחי קריירה.
הרולד היה נטול פשרות ביושרו מבחינה אינטלקטואלית, מסור לניהול חיים המבוססים על עקרונות ומונע על ידי מצפן מוסרי חזק שהשפיע על כל מי שפגש. ביטויה התגלה בכל תחומי חייו – משפחתו, הגותו ההומניסטית…, סלידתו מהקפיטליזם ומחויבותו לחשיפת שחיתות מוסדית, צמצום החופש האקדמי, טיוח מחקרים מופרכים, אובדן זכויות אדם ופגיעה ברווחת בעלי חיים…"(מקור 1)
כשעוברים על עבודתו וראיונותיו של הילמן, נראה שהאמור לעיל הוא תיאור מדויק של האיש. תוכלו לקרוא את סיפור הקריירה שלו ואת המחירים ששילם על יושרו האינטלקטואלי.(מקור 2)
הילמן היה מדען נדיר, בכך שמוקד הקריירה שלו לא התרכז בלהוכיח או לגלות משהו חדשני. במקום זאת, הוא התמקד בבדיקת עובדות ובביקורת על מה שנחשב לעובדות מוכחות, שהיוו את היסודות של תחום הביולוגיה שבחר. עובדות במובן זה שהן רעיונות הנלמדים כאמיתוֹת בדוקות הניתנות לאימות על ידי מורים, מרצים ומחברים של ספרים ללימוד ביולוגיה עד עצם היום הזה.
כשקוראים את ספריו של הילמן, "התא החי" (1980) ו"ודאות וחוסר ודאות בטכניקות ביוכימיות" (1972), מקבלים מושג טוב על יושרו האינטלקטואלי הבלתי מתפשר של הילמן. עם זאת, ככל שאתה עובר על חלק ניכר מעבודתו של הילמן אתה מתחיל להבין שהוכחת יושר אינטלקטואלי הייתה הנושא הבסיסי של כל עבודתו המדעית, המוקד האמיתי של הקריירה שלו.
יושר אינטלקטואלי בניסויים מדעיים בכל ענף מחייב יישום שיטתי ובתום לב של השיטה המדעית.
משמעות הדבר היא זיהוי כל ההנחות, לרבות אלה הקשורות להליכים שננקטו לביצוע הניסוי – דהיינו, זיהוי המגבלות של כל מכשיר, תכשיר ו/או נוהל לביצוע הניסוי. זה גם אומר להפנות את תשומת הלב לאופן שבו מגבלות אלה משפיעות על המסקנות הסופיות שהוסקו מהניסוי.
זה אומר ביצוע ניסויי ביקורת בשלבים המתאימים של הניסוי, הכוללים ניסויים לאימות הדיוק של הציוד בו נעשה שימוש. זה אומר לפרסם את התוצאות של ניסויי הביקורת האלה בכל מקרה ובמיוחד כאשר הם מטילים ספק בתקפות ההשערה הראשונית.(מקור 3)
היישום של השיטה המדעית על ידי הילמן באופן שתואר לעיל לחקר התאים והמבנים הפנימיים שלהם, הוביל את הילמן לביקורת נוקבת על הביולוגיה המודרנית.
כפי שנאמר על ידי ד"ר טום קוואן, עבודתו של הילמן הראתה כי רבות מהעובדות הבסיסיות של הביולוגיה רחוקות מאוד מלהיות באמת עובדות, עד כדי כך שיש צורך להעריך מחדש את כל התחום של ביולוגיית התא המודרנית [ביולוגיה מולקולרית], שהיא התשתית של הרפואה המודרנית.(מקור 4) ממה שקראתי על עבודתו של הילמן, קוואן לא הגזים.
למה שיהיה לך בכלל אכפת מהתחום של ביולוגית התא המודרנית?
ובכן, זאת מאחר שהאופן שבו אנו חושבים על התא עומד בבסיס האופן שבו אנו חושבים שיישויות חיות פועלות. התא נחשב לדבר החי הקטן ביותר שממנו מורכב כל יצור חי – יחידת החיים הבסיסית. נאמר לנו כי תאים מפעילים את כל התהליכים הדרושים כדי לקיים חיים, כגון סינתזה (יצור) של חלבונים. ונאמר גם כי תאים נובעים רק מתאים אחרים.
נאמר כי הדנ"א שלנו, וכך גם הגֶנים שלנו שוכנים בתוך התאים שלנו, והם משוכפלים ומועברים לכל תא חדש שנוצר באמצעות חלוקה התא. כך, נאמר לנו, הגֶנים ותאים הם האחראים למאפיינים התורשתיים שלנו. גֶנים אמורים גם להיות אחראים לרגישות של אדם או בעלי חיים למחלות מסוימות. מהן, אם כן, הביקורות המשמעותיות הללו על [מחקר] הביולוגיה של התא?
למרבית עבודתו של הילמן יש השלכות מרחיקות לכת על הביולוגיה והרפואה המודרנית, אך בהתחשב בכך שמאמר זה נרחב דיו כדי לכסות בפירוט רק היבט אחד של עבודתו, ניסיתי לבחור היבט שעשוי להיות המעניין ביותר ובעל ההשלכות מרחיקות הלכת ביותר – ביקורתו של הילמן לגבי מבנה התא החי וכיצד היא משפיעה על הבנתנו את סינתזת החלבונים. באשר לשאר עבודתו, אני מקווה שמאמר זה יעורר מספיק עניין כדי שהקורא ירצה להתבונן בעצמו בעבודותיו האחרות של הילמן.
לפני שנצלול פנימה, חשוב להבין מראש למה הכוונה במושג "תא חי". כאשר הילמן מתייחס ל"תא החי" הוא מתכוון לתא [של גוף] שעדיין נמצא בתוך אדם או חיה חיים (in vivo) ולא כזה שהופרד מהיצור החי (למעשה הופרד ממקור החיים שלו) וטופל לצורך המחקר.
בעשותו הבחנה זו, הילמן מדגיש את העובדה שאחת ההנחות המשמעותיות ביותר של הביולוגים היא שהפרדת דגימת של רקמות או תאים ממקור החיים שלהם [הגוף החי] אינה משפיעה על [מצב] הדגימה או על הכימיה שלה, וכי בנוסף, במקרים בהם יש להרוג את בעל החיים, [ביולוגים מניחים ש]הלחץ של הטיפול בבעל החיים וההרג שלו אינם משפיעים על תוצאות הניסוי.
בסרטון זה מובא ההסבר של הרפואה החדשה, מדוע ניסויים בבע"ח הם נטולי ערך מחקרי.
הסרטון אינו חלק מהמאמר המקורי
2. הילמן ומבנים של התא-הכללי
הערכתו המחודשת של הילמן את מבנה התא החי החלה ברצינות כאשר הוא ועמיתו, פיטר סארטורי, נתקלו בכמה סטיות (אנומליות) חשובות בין המבנה וההתנהגות של התא כפי שהם נראים במיקרוסקופ אור ברקמה לא-מקובעת, מצד אחד, לבין המבנה כפי שהוסק ממיקרוסקופ אלקטרונים ברקמה מקובעת, מצד שני.
א. מיקרוסקופ אור ומיקרוסקופ אלקטרונים
מיקרוסקופ אור או מיקרוסקופ אופטי ממקד קרן אור נראה (פוטונים) דרך דגימה כדי להפיק תמונה ברזולוציה של 200-250 ננומטר (nm). התמונה שנוצרת כך מורכבת מפוטונים שלא נעצרו בדגימה וכאלה שפוזרו [עברו דיפרקציה] על ידי הדגימה.
תמונה זו מועברת דרך עדשה אחת או שתיים כדי להגדיל את התמונה לצפייה. זרם הפוטונים יכול להיות מכוון אל הדגימה בכל זווית, כלומר ניתן לבחור זוית טבעית של הדגימה כאילו הדגימה הנבדקת נמצאת בידו של הבודק.
מיקרוסקופ אור יכול לצפות בדגימות חיות, מתות או מקובעות ולרשותו מגוון רחב של שיטות הכנה וצפייה (שדה בהיר, ניגודיות פאזה וכו '), אך במאמר זה נתייחס רק לתצפית על דגימות חיות תחת מיקרוסקופ אור.
בעת התבוננות בדגימות חיות (לא צבועות ולא מקובעות) הדגימה מונחת רק על שקופית מיקרוסקופ לפני שהיא מונחת במיקרוסקופ לצפייה.
מיקרוסקופ אלקטרונים משתמש באלומה ממוקדת של אלקטרונים כדי "לצלם" את הדגימה ברזולוציה של 1.5 עד 2 ננומטר. עם זאת, התהליך שנעשה כדי לייצר מיקרוגרף [צילום] אלקטרונים הוא הרבה יותר מסובך מזה הנדרש במיקרוסקופ אור.
ראשית הרקמה מקובעת (מומתת) בפורמלין, אשר מפסיק את חילוף החומרים ומעכב פעילות אנזימים. בעיקרון, הרקמה מטופלת כדי למנוע ממנה להירקב לפני שהיא נצפית.
לאחר מכן, מכיוון שהרקמה לא תחזיק מעמד בתוך ואקום חזק ביחד עם הפצצת אלקטרונים וקרינת רנטגן, יש לצפות את הרקמה, או לצבוע אותה, על ידי משקעים של מלחי מתכת כבדים כגון אוסמיום, עופרת, טונגסטן, מנגן וכו '.
הרקמה הצבועה מיובשת לאחר מכן בריכוז הולך וגובר של אלכוהול (אשר מכווץ את הרקמה). לאחר מכן מוצא האלכוהול מהרקמה באמצעות ממיס שומן (פרופילן אוקסיד), ולאחר מכן הרקמה מוספגת ומוטמעת בתוך שרף אפוקסי (פלסטיק), אשר מושאר ולאחר מכן למספר ימים כדי להתקשות.
חלקים או פרוסות בעובי עשירית המילימטר, או פחות, נחתכים באמצעות כלי חיתוך מיוחד הנקרא מיקרוטום. שימו לב כי למיקרוסקופיסטים [מפעילי המיקרוסקופ] אין מושג על כיוון להב המיקרוטום ביחס לתא ולמבנים הפנימיים שלו בעת שהם חותכים דגימות.
דגימות דקות להפליא אלה ממוקמות בתוך תא הצפייה של המיקרוסקופ, שם נשאב כמעט כל האוויר החוצה כדי להשאיר את הדגימה בוואקום גבוה, מה שמכונה אוורור (venting).
קרן אלקטרונים במתח שבין 1,000 וולט ל- 3,000,000 וולט מכוונת אל הדגימה, כמות הוולטים תלויה בהגדלה הרצויה. ככל שהמתח גבוה יותר, כך ההגדלה גבוהה יותר.
אותם אלקטרונים שאינם נחסמים על ידי המתכות הכבדות, עוברים דרך הדגימה ופוגעים במסך זרחני. המיקרוסקופיסטים בוחרים שדה (אזור) מסוים במסך ומתקרבים עד שניתן להבחין בתכונות שהם רוצים להראות. לאחר מכן ניתן לשפר את התמונה ולצלם תמונות, שחלקן נבחרות כראיות.
יש הטוענים כי טכניקות הקפאה עמוקה מהירה ומודרנית יותר, בהן הדגימות מוקפאות ב -150 מעלות או יותר, יכולות להתבצע מבלי לגרום להתייבשות נוזלי התא כמו שקורה עם אלכוהול.
הילמן אומר כי ללא ביצוע ניסויי בקרה נאותים הדבר מעולם לא הודגם. יתר על כן, הוא חולק על כך שזה יכול להיות המצב, בהתחשב בכך שכאשר משהו קפוא לטמפרטורות נמוכות מאוד, מים יוצאים מכל תמיסה שבה הם נמצאים בצורה של גבישי קרח, ובכך משאירים את כל המרכיבים האחרים של התמיסה מיובשים.
כמו כן, חיוני להבין כי בנוסף לשלבים המייבשים את הדגימה במהלך ההכנה, הדגימה מיובשת עוד יותר כאשר היא נחשפת להפצצה של קרן האלקטרונים. הסיבה לכך היא שכאשר קרן האלקטרונים פוגעת בדגימה, היא מחממת את השילוב רקמה-מתכת וגורמת לטמפרטורת הדגימה לעלות במאות מעלות.
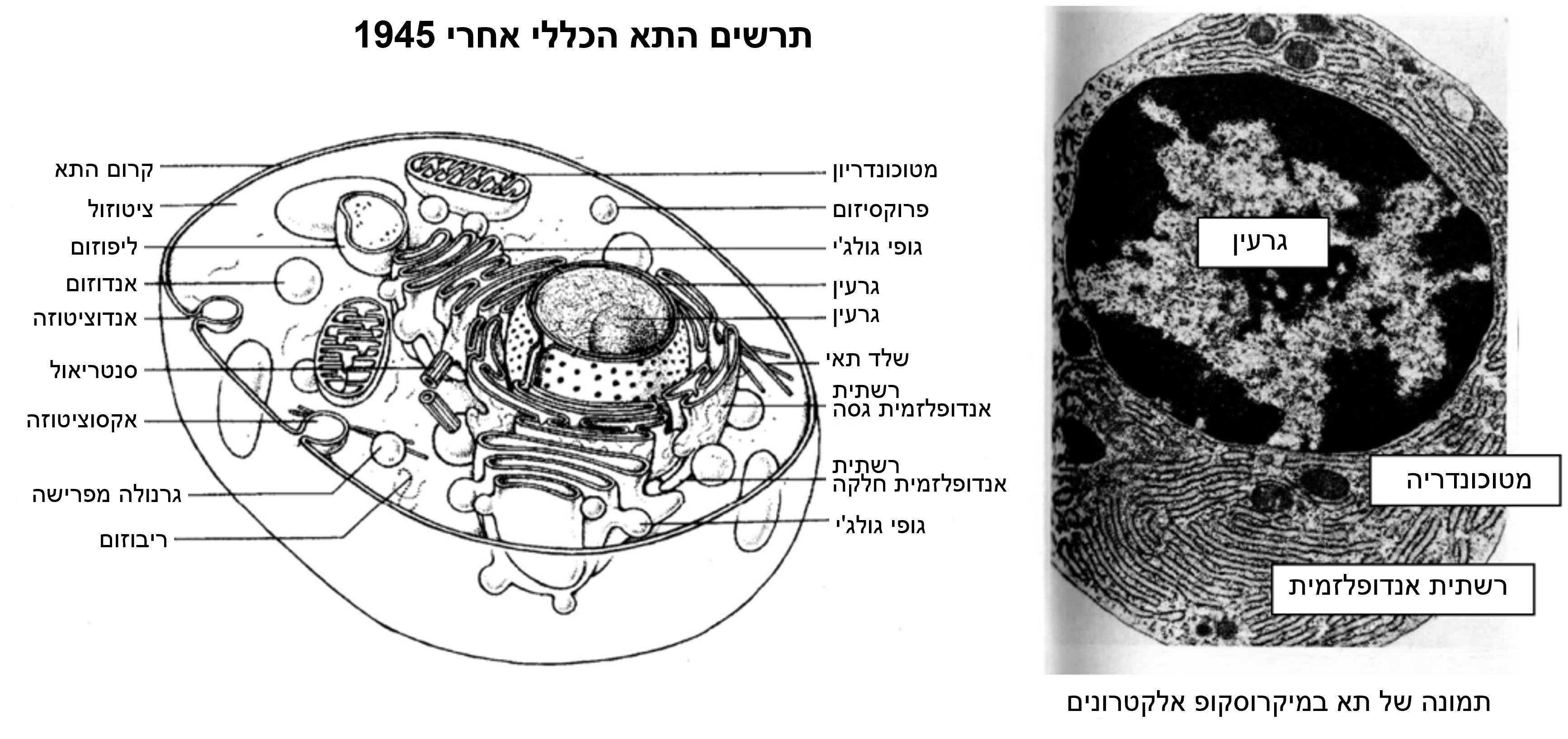
ב. התא-הכללי
בהמשך לחקירותיהם, הילמן וסארטורי הגיעו למסקנה כי המודל המקובל של התא (מודל התא-הכללי) אינו אפשרי מסיבות גיאומטריות וביולוגיות, וכי מבנים פנימיים אלה (הידועים גם בשם אברונים) שהוסקו על ידי מיקרוסקופ אלקטרונים הם תוצרי לוואי של השיטות המשמשות להכנת הדגימה לצפייה.
עד תחילת שנות ה- 1940, לפני השימוש במיקרוסקופ אלקטרונים לחקר תאים, הוסכם שכל התאים החיים הם ישויות עגולות או כדוריות המורכבות מקרום תא חיצוני התוחם את הכדור, עם מיטוכונדריה, גרעין, ציטופלסמה והקרום של הגרעין.

בעקבות התחלת השימוש במיקרוסקופ האלקטרונים, האברונים הבאים, שניתן להוכיח שהם [תוצרים] מלאכותיים, נוספו למודל התא הכללי:(מקור 5)
- קרומי התא, הגרעין והמיטוכונדריה מופיעים כעת כולם כשני קווים עם רווחים ביניהם, מראה זה נקרא "קרום התא" (unit membrane’) והוא עומד בניגוד לקו היחיד שהיה מקובל לפני 1940;
- "רשתית אנדופלסמית" (ר"א) הנתפסת כמבנה תלת ממדי בציטופלסמה (הג'ל הנוזלי הממלא את קרום התא, חיצונית לגרעין התא);
- "ריבוזומים" המרפדים את הרשתית האנדופלסמית;
- "נקבוביות הגרעין" הן חורים המופיעים בקרום הגרעין;
- "ליזוזום" הוא מבנה תאי הקשור לקרום התא ומצוי בציטופלסמה, המכיל אנזימים מעכלים;
- "Cristae" מעין קפלים שתוארו כנמצאים במיטוכונדריה.
3. "קרום התא [ממברנה]"
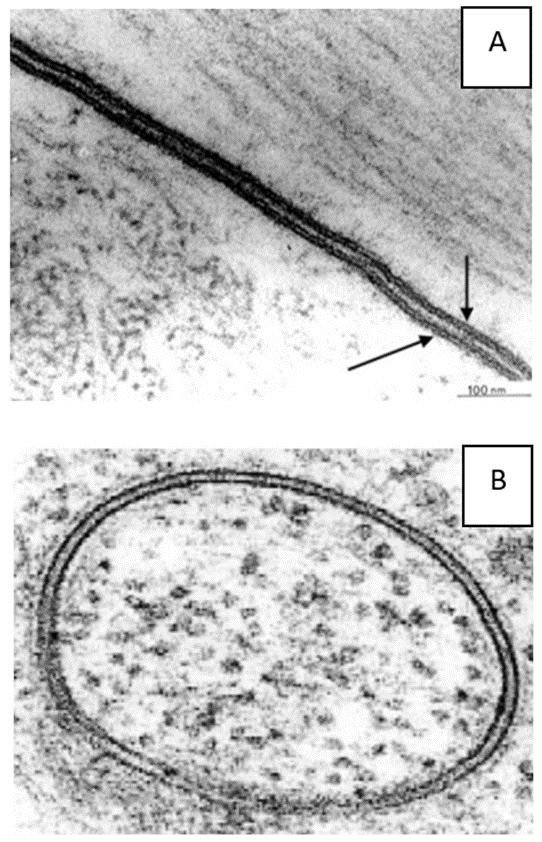
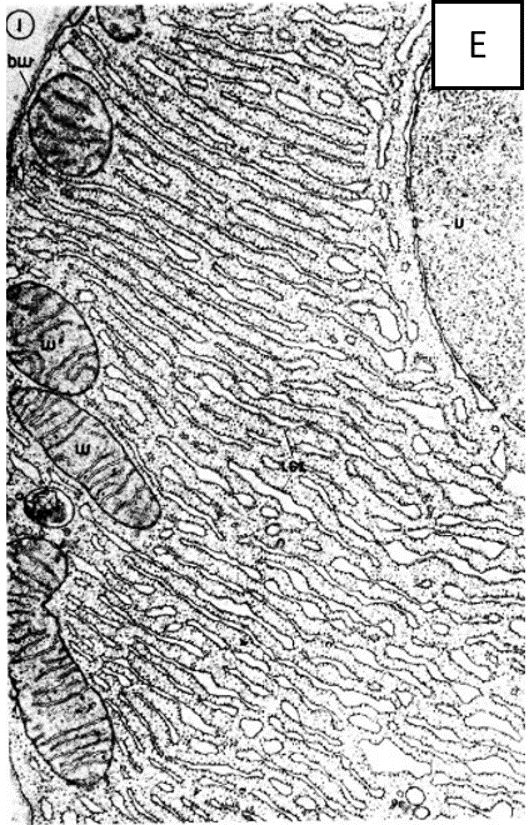
המעטפות של התא, גרעין התא והמיטוכונדריה מופיעים כולם במיקרוסקופ אלקטרונים כשני קווים עם רווח ביניהם, מבנה דמוי כריך זה קיבל את השם "קרום התא". (מקור 6) ראו מיקרוגרפים אלקטרונים [צילומי מיקרוסקופ האלקטרונים] A ו-B.
הילמן וסארטורי טוענים למספר בעיות במסקנות לגבי מבנה "קרום התא", להלן הסוגיות הברורות ביותר.
א. טיעונים גיאומטריים
כשמתבוננים במיקרוגרפים אלקטרוניים המתארים קרומים, שני הקווים החיצוניים של ה"כריך" נראים תמיד מרווחים זה מזה באופן אחיד להפליא.
הבעיה כאן היא שכאשר חותכים מקטעים של כדורים קונצנטריים (בעלי מרכז משותף. חשבו על פריסת שכבות של בצל עגול לחלוטין), אנו מצפים לראות שהמרווח הנראה לעין בין השכבות ישתנה בהתאם לקרבת הפרוסה למרכז הכדור או לקצהו.
בהתחשב בכך שכל המבנים הממברניים בדגימה יהיו אקראיים, ושהמיקרוסקופיסט אינו יכול לדעת מראש מאיזו זווית יחתוך אותם המיקרוטום, הרי שגם אם לשכבות השונות היה בדיוק אותו עובי [בתא החי], הן היו צריכות להופיע במגוון עוביים בתוך סדרה של חתכים (ראו איור 1).
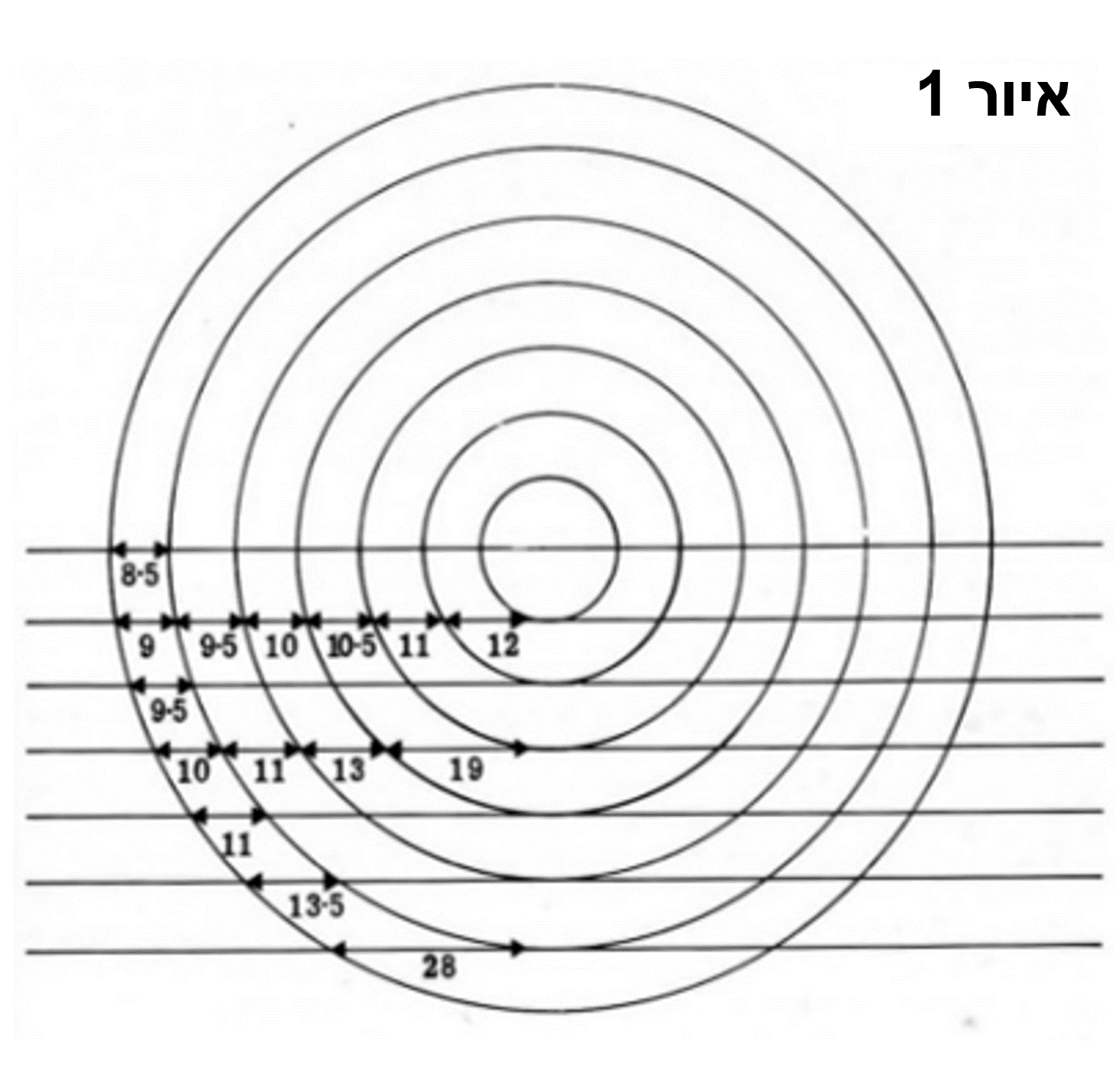
במילים אחרות, בקטעים שונים שצולמו מאותה ממברנה, עובי השכבות צריך להשתנות וגם כמות השכבות הנראות צריכה להשתנות, כך שצריכים להיות מקרים שבהם רק מחצית מהשכבות גלויות ואחרות שבהן הרווח באמצע אינו נראה כלל. ראו איור 2 להלן.
הסיבה לכך היא שהאופן שבו שכבות הממברנות מופיעות בחתך חייב להיות תלוי בהכרח במקום שבו נחתך הכריך. האפשרויות הן: דרך מרכז התא (שלוש שכבות תיראנה במרחק שווה וגלויות לצופה – משמאל), דרך חלק מעובי הקרום בלבד, קרוב לחלקו החיצוני של התא, בין שתי שכבות הממברנה (שכבה אחת וחלק מהרווח בין השכבות יהיו גלויים – מימין) או לגמרי לצד בלי הממברנה (ללא רווח נראה לעין מנקודת המבט של הצופה – באמצע).
העובדה שבמיקרוגרפים אלקטרוניים שכבות ממברנות נראות כשומרות באופן עקבי על רוחבן ללא קשר לכיוון החתך, פירושה שכל המבנים הנראים בקטע מסוים היו צריכים להיות: א) במיקום מרכזי וסימטרי לחלוטין זה לזה בזמן יצירת החתך, וגם ב) להחתך בדיוק דרך מרכז המבנה שלהם באותו הזמן. זאת למרות שכל המבנים הנראים לעין שונים מאוד בגודלם ומיקומם.
סדרה כזו של נסיבות הקשורות זו בזו היא אבסורדית ומאוד לא סבירה. מה שהרבה יותר סביר שקורה הוא, שקרום ה"יחידה" הוא דו-ממדי וכי השכבה הכפולה (שהיא המרמזת על שלושה ממדים) מופיעה רק לאחר החיתוך, בעת שמכינים את הדגימה לצפייה או במהלך הפצצת האלקטרונים עצמה.
בעיה נוספת עם הופעתם של הקרום "הכפול" של התא היא העובדה שמעטפות התא, הגרעין והמיטוכונדריה נתפסות כמעוגלות, בהתחשב בכך שהן מקיפות דברים. משמעות הדבר היא שבכל חתך בודד נתון, נוכל בהכרח למצוא מקרים שבהם הרווח בין שכבות הקרום אינו נראה כלל לעין מפני ששתי השכבות החיצוניות נראות חופפות ומכסות על המרווח – לדוגמה כאשר השכבות הכפולות נעלמות מעבר לפינה, כביכול.
במקום זאת, לעתים קרובות מוצגות בפנינו שכבות קרום כפול השומרות על מרחק אחיד בניהן סביב כל ההיקף.
ב. טיעונים ביולוגיים והפרדה תת-תאית
כשמדובר בביוכימיה, מקובל לחשוב כי שכבות "קרום התא" הן שתי שכבות חלבון שבניהן שכבת שומנים (ליפידים).(מקור 7)
בנקודה זו חשוב להבין מהי הפרדה תת-תאית (subcellular fractionation) וכיצד ביוכימאים וביולוגים מולקולרים משתמשים בה, כדי לקבוע את מיקומן של מקרו-מולקולות שונות (כימיה) בתוך התא החי וכדי לבחון את המבנה (מורפולוגיה) של אברונים בודדים.
הפרדה תת-תאית הוא תהליך שתוכנן כדי להפריד תאים למרכיביהם השונים (אברונים) תוך השארת האברונים שלמים במידה רבה ושמירה על תפקודם האינדיבידואלי.
חשוב לציין כי לפי האמונה, ההליכים הללו אינם משנים את המאפיינים של האברונים, ושלאחר הפרדתם מהתא החי ואחד מהשני, האברונים שומרים על תכונותיהם באותה המידה כאילו הם עדיין בתוך יצור חי שלם.
השלב הראשון בהפרדה תת-תאית דורש שהדגימה תעבור תהליך האחדה [הפיכת תערובת להומוגנית] (על ידי טחינה, מעיכה קיצוץ או לחיצה) בתוך תמיסת חיץ לא ביולוגית (תמיסה על בסיס מים וייחס משתנה של חומצה ובסיס).
ההאחדה מבטיחה שקרום התא יישבר ותכולת התא תשוחרר לתמיסת החיץ. תמיסת החיץ מגנה על האברונים על ידי שמירה על רמת ה-PH הרצויה של התמיסה ובכך מעכבת פעילות אנזימתית (עוצרת תהליכי ריקבון ותגובות כימיות).(מקור 8)
לאחר מכן מוסיפים סוכרוז [סוכר לבן] לנוזל האחוד (הומוגנט) ואז מסחררים אותו בצנטריפוגה. תהליך זה מורכב מהכנסת הדגימות לצנטריפוגה – מכונה שנועדה לסובב דגימות נוזל במבחנה במהירויות גבוהות – כך שהכוח הצנטריפוגלי [הכוח הדוחה מן המרכז בתנועה סיבובית מהירה] גורם להצטברות המשקעים הגדולים או הכבדים בצורת מעין גליל או גוש בתחתית המבחנה, בעוד החלקיקים הקטנים או הקלים יותר צפים למעלה.
סוכרוז בתמיסה משמש כמדיום מכיוון שצפיפותו אינה מושפעת מהסחרור בצנטריפוגה ולכן הוא אינו מפריע להפרדת האברונים לשכבות משקע. ניתן לצפות בסרטון של תהליך הצנטריפוגה.(מקור 9)
המהירות, משך הזמן ומספר הפעמים שהדגימה עוברת צנטריפוגה תלויים באברונים אותם מבקשים לחקור.
באופן כללי, האמונה היא כי אברונים קטנים יותר ידרשו יותר סבבי צנטריפוגה מאשר אברונים גדולים יותר, וכל סבב צריך להיות במהירויות גבוהות יותר ולפרקי זמן ארוכים יותר.
מהירות נמוכה מכפיפה את הנוזל האחוד (הומוגנט) לכוח גדול פי 1000 מזה של כוח הכבידה למשך 10 דקות, ומהירות גבוהה מאוד מכפיפה את הנוזל האחוד לכוח גדול פי 150,000 מזה של כוח הכבידה למשך 3 שעות.(מקור 10)
המשקעים המופרדים, המכונים שברים, אשר נדחפים לתחתית לאחר כל סבב של צנטריפוגה, נצפים תחת מיקרוסקופ אלקטרונים כדי לקבוע את טוהרם. אם נקבע שהשבר אינו טהור מספיק (יש יותר פסולת תאית ואברונים אחרים מהאברון הרצוי), ניתן יהיה לסחררו שוב בצנטריפוגה.
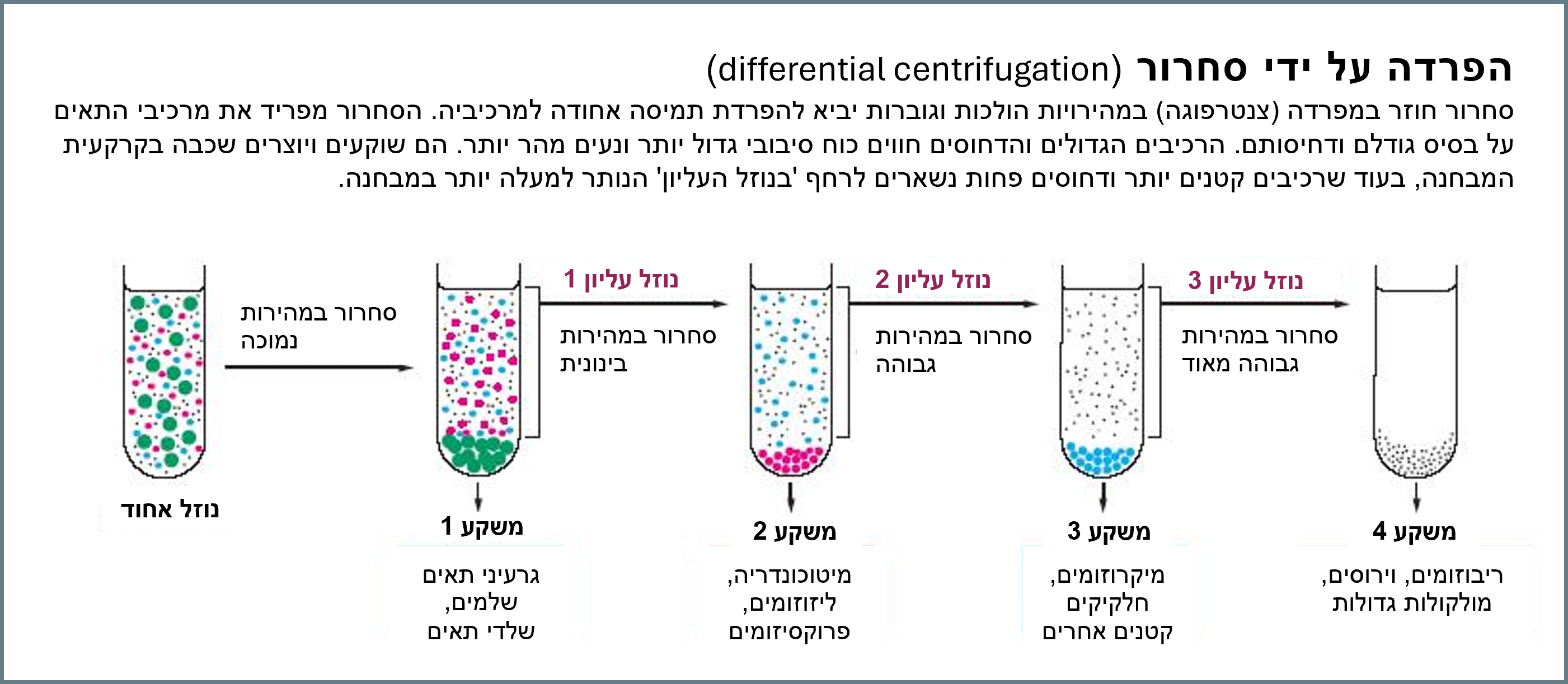
הביולוגים מאמינים שהתהליך לא משנה את הדגימה הנבדקת
המקרו-מולקולות, כגון חלבונים או חומצות גרעין, הנמצאות בכל שבר, מיוחסות לאחר מכן לאברונים שהיו אמורים להיפרד בצפיפויות אלו. שימו לב שמקרו-מולקולות אלה מיוחסות לא רק לשברים המופרדים, אלא גם לאברונים שלפי האמונה נמצאים בתא החי.
הילמן וסארטורי מטילים ספק בכך שקרומים מורכבים מליפידים וחלבונים, שכן הם מתייחסים לראיות מהאנליזה הכימית של שברים תת-תאיים כמפוקפקות מדי, עד לביצוע ניסויי ביקורת רלוונטיים. ללא ניסויי ביקורת, לא ניתן לדעת מהי ההשפעה של תהליכי ההאחדה, הצנטריפוגה והתוספים הכימיים הנדרשים, על הדגימה ועל הביוכימיה שלה.
ההליך כולו נשען גם על ההנחה שכל אברון תורם רק בדרך אחת ספציפית לתהליכים הביוכימיים של התא (הנחת מידור).(מקור 11)
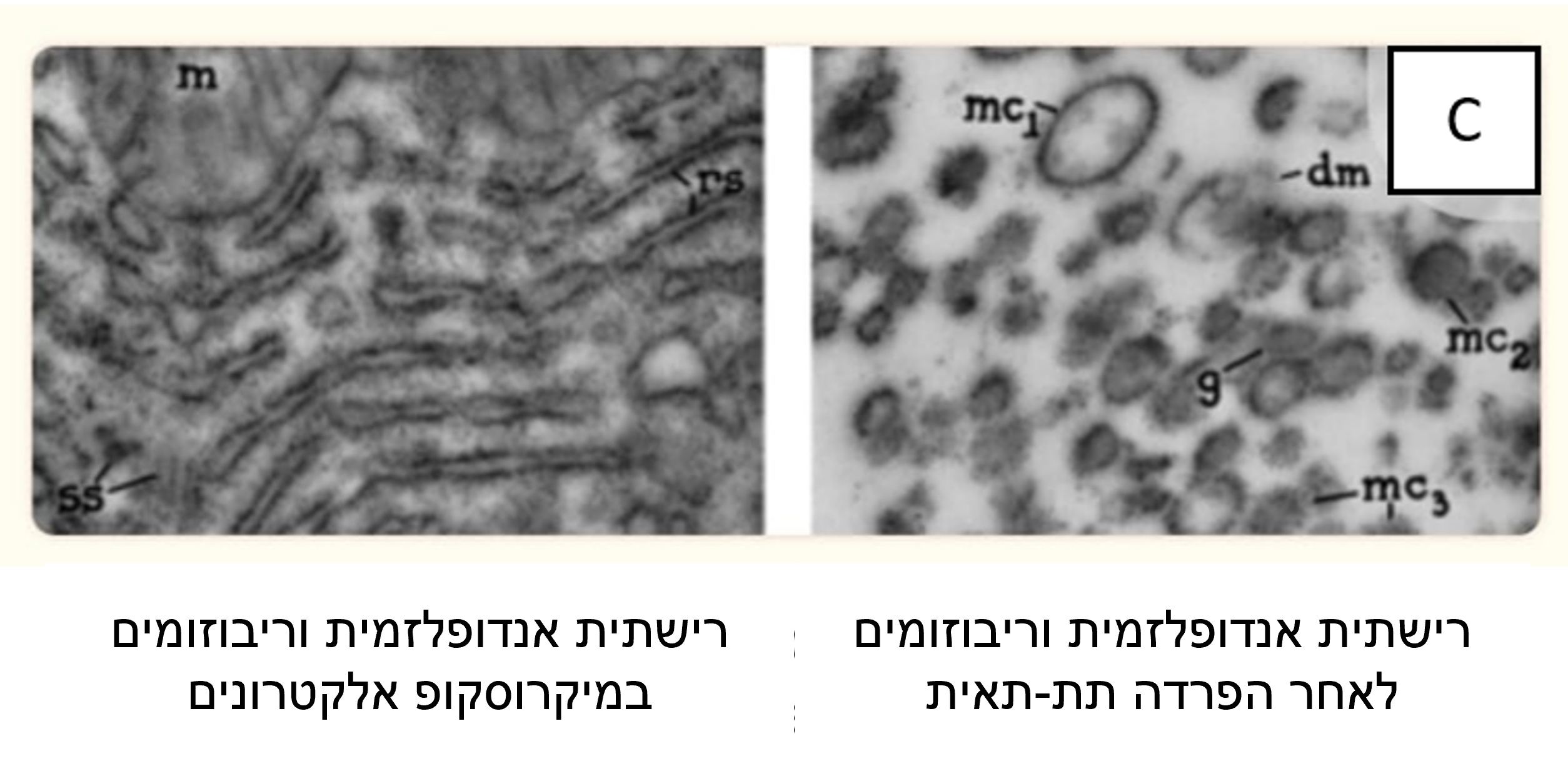
ראוי גם לציין כי בעוד את המיטוכונדריה וגרעיני התא קל למדי לזהות בתוך השברים, הרי שזהוי הקרומים (ממברנות), הריבוזומים והרשתית האנדופלסמית קשה הרבה יותר.
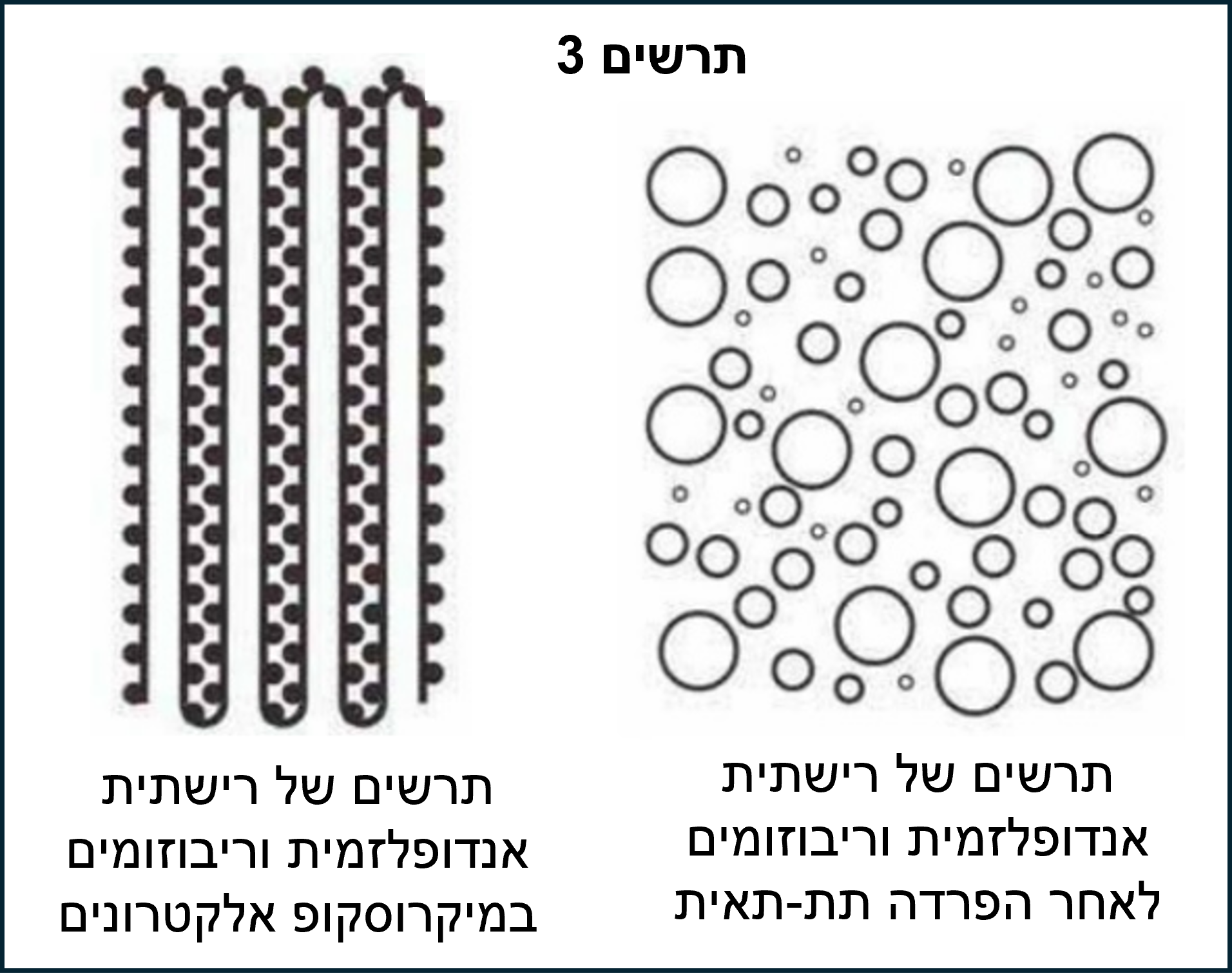
הסיבה לכך היא שריבוזומים הם נטולי צורה מוגדרת בבירור והרשתית האנדופלסמית והקרומים אמורים להופיע כעיגולים או אליפסות בשברים. החלקיקים המעגליים הנראים ברשתית האנדופלסמית, בריבוזום ובשברי הקרום ידועים בשם "מיקרוזומים". ראו מיקרוגרף אלקטרונים C ותרשים 3 .
הדבר גם משמעותי אם לוקחים בחשבון שבתהליך ההפרדה התת-תאית, ההצדקה לכל הליך ומשכו, ולכל תוסף ווריכוזו, מבוססת אך ורק על ההתאמה, כביכול, במיקרוסקופ אלקטרונים, בין המבנים המופרדים לאלה שברקמה השלמה.
סיבה נוספת לכך שהילמן וסארטורי מפקפקים בכך שהקרומי המעטפת מורכבים משומנים וחלבונים היא מכיוון שבמהלך קיבוע רקמות משתמשים בממסים (אתנול, אצטון) הידועים כממיסי שומנים ולאחריהם קשה להאמין שניתן לראות שומנים במיקרוגרף האלקטרונים.(מקור 12)
שימו לב כי הנושאים לעיל, הנוגעים למראה ולביוכימיה של "קרום התא", רלוונטיים גם לרשתית האנדופלסמית שתידון מיד להלן.
4. "רשתית אנדופלסמית" (ר"א)
רשתית אנדופלזמית(מקור 13) נחשבת בעיני רבים לאחד הממצאים החשובים ביותר של מיקרוסקופיית אלקטרונים. דיאגרמות שלה נמצאות בכל ספרי הלימוד של מדעי החיים, המשמשים תלמידי בית ספר כמו גם סטודנטים באוניברסיטה, סטודנטים לתואר שני ועובדי מחקר מקצועיים.
זהו האברון הגדול ביותר בתא ונאמר כי הוא משמש אתר מרכזי של סינתזת [יצור] חלבונים, הובלת חלבונים וקיפול חלבונים.(מקור 14)
האמונה היא שאברון זה נמצא בכל תאי הצמחים ובעלי החיים כרשת תלת ממדית או כמערכות של שקים שטוחים, או בועיות, החודרות דרך חלק או כל נפח הציטופלסמה. נראה גם שיש לו שתי שכבות עם רווח ביניהן, כמו במקרה של "קרום התא". ראו מיקרוגרפים אלקטרונים D ו-E.
הר"א מיוצגת לרוב כמחוברת לקרום התא ולקרום הגרעין.(מקור 15) היא נראית לעיתים כמרופדת ב"ריבוזומים" שהם גרגירים, אשר מאמינים שהם מורכבים בעיקר מחומצה ריבונוקלאית (רנ"א) ומעורבים גם בסינתזה של חלבונים.
האמונה היא, כמובן, כי הר"א קיימת בתאים חיים כפי שהיא מופיעה במיקרוגרפים אלקטרונים, דבר המשתמע מההשקפה שהן הר"א והן ה"ריבוזומים" שמרפדים אותה, מעורבים בסינתזה של חלבון.
לא רק שקיום ר"א בתאים התקבל באופן אוניברסלי בקרב מורפולוגים, אלא שהביוכימאים יצרו הפרדות תת-תאיות אותן הם מזהים כשייכים לר"א (הם מכונים "מיקרוזומים", כפי שהוסבר לעיל) והם חקרו את הביוכימיה שלה בהרחבה. תפקודה נמצא אפוא במתאם עם המבנה שלה, דבר הנחשב כראיה מאשרת לקיומה בתא.
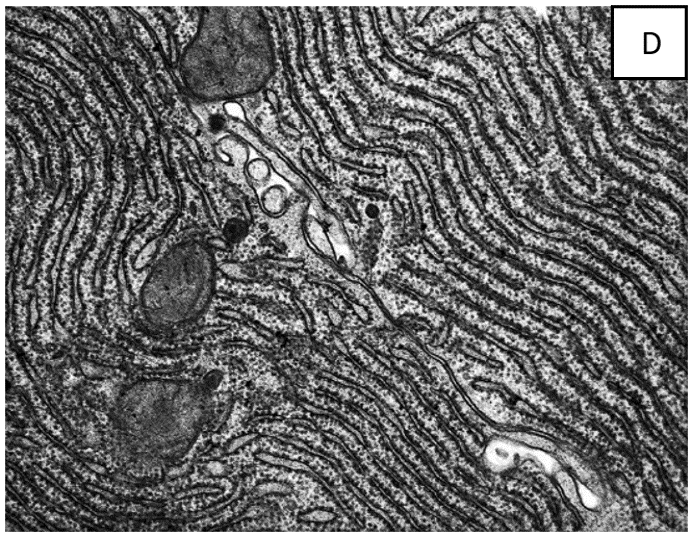
הבעיה הגדולה ביותר של הילמן וסארטורי עם ההסבר לעיל היא שבכל מיקרוגרף אלקטרונים המסופק כראיה למיון בתאים, הר"א נראית תמיד רק באותו מישור כמו התמונה, בחתך רוחבי.
הבעיה עם זה היא, שאם הר"א היתה רשת או סדרה של שקיות שטוחות, אפשר היה לצפות לראות אותה בחתך מכל מיני כיוונים ולא רק מאחד. אם [בעת החיתוך] הר"א נמצאת במנח אקראי בתוך התא, הסכין היה צריך לחתוך אותה בצורה אקראית. בנוסף, בסדרה של חתכים כאלה הר"א היתה צריכה להראות חתוכה מסדרה של כיוונים.
במילים אחרות, אם הר"א היא מעין רשת העשויה קרום בדומה לקרום התא, מדוע אנו רואים תמיד רק חתך מכיוון אחד שלו בתאים? איפה התמונות של חתכים מכל הכיוונים השונים שהיינו מצפים לראות? זה די בלתי אפשרי להעלות על הדעת אובייקט תלת-מימדי, אשר נראה תמיד אותו הדבר בשני ממדים, כאשר חותכים אותו מכיוונים שונים.
לדוגמה, מדוע אנו רואים את הר"א על מיקרוגרפים אלקטרונים רק [בכיוונים ניצבים] כמו בתמונה 4a או 4b, אך אף פעם לא [בכיוונים משופעים] כמו תמונות 5a עד 5f:
המיקרוסקופיסטים האלקטרונים טוענים כי אין תמונות של ר"א מכיוונים אחרים, מפני שמפרסמי המאמרים בוחרים רק את התמונות בהן אפשר לראות את הר"א בבירור.


משום מה, הרשתית האנדופלזמית אף פעם לא מופיעה במנח אלכסוני
הילמן וסארטורי אומרים שזה שטויות, לא רק שהם עצמם צפו בתאים תחת מיקרוסקופ אלקטרונים, אלא שהחקירות שלהם את האנומליה הזו הובילו אותם לבחון מיקרוגרפים אלקטרונים של תאים שלמים רבים ככל שיכלו למצוא, אלפי מיקרוגרפים אלקטרונים של תאים, כאלה שלא פורסמו וכאלה שכן פורסמו. כולל כאלה שבהם המחברים הפנו את תשומת הלב למבנים שאינם הר"א.
ראוי גם לציין כי בניגוד גמור לר"א – מיטוכונדריה דוקא כן מופיעים במיקרוגרפים אלקטרונים בכל מנח וצורה שבין גליל למעגל, בהתאם למישור החתך. ניתן לראות מטוכונדריה בכל מיקרוגרף אלקטרונים שנבחר באופן אקראי, והתמונה של מבנים תאיים אלה מהווה מודל בקרה מצוין לתצפיות על הר"א.
ראו מיקרוגרף אלקטרונים F, למשל. העיגולים הכהים הקטנים והאליפסות בגדלים שונים הם המיטוכונדריה בכיוונים שונים במישור החתך. כמובן, מיטוכונדריה ניתן לראות גם על ידי מיקרוסקופ אור וקיומם אינו מוטל בספק.

סיבה שנייה לכך שהר"א נראית כתופעת לוואי מלאכותית היא העובדה שבמיקרוגרפים היא נראית עבה יותר מאשר התאים ומממברנות הגרעין, שאותם כן ניתן לזהות במיקרוסקופ אור. בנוסף, ההשקפה הנוכחית לפיה "ריבוזומים" מרפדים את הר"א בתאים מסוימים, צריכה היתה לגרום לה להראות עבה עוד יותר. מדוע, אם כך, לא ניתן לזהות את הר"א במיקרוסקופ האור?
קושי נוסף לגבי קיומה של הר"א בתא החי הוא שהיא אמורה להיות מחוברת ל[מעטפת] התא [מבחוץ] ולקרום גרעין התא [מבפנים]. כיצד יכול להתרחש סיבוב של הגרעין אם הר"א מחוברת הן לתא והן לקרום הגרעין, שכביכול מלא בנקבוביות? סיבוב הגרעין מתייחס לעובדה שתחת מיקרוסקופ אור ניתן לצפות בעיבויים אקראיים (בליטות) לאורך קרום הגרעין, אשר מסתובבים עם הגרעין. תופעה זו, יחד עם תנועות בין-תאיות אחרות, נצפתה בתאים חיים מאז 1954 והיא אחד הקריטריונים המשמשים מיקרוסקופיסטים כדי לדעת אם התאים בדגימת הרקמה עדיין חיים.
הנקודה כאן היא שאם הר"א מחוברת לשני הקרומים ואחד מהם, קרום הגרעין, מסתובב כל הזמן, אזי א) איך הר"א עצמה לא מסתבכת, ב) איך המיטוכונדריה, הגרגירים ופסולת התא לא נתפסים בר"א, ג) איך חלקיקים שהוכנסו לציטופלסמה מתפזרים באופן שווה ולכל הכיוונים, כפי שניתן לראותם עושים במיקרוסקופ אור?
אם תעלה טענה כי הר"א אינה מחוברת לגרעין המסתובב, אזי יש להודות כי הר"א לא יכולה לשמש כערוץ לייצוא החלבונים, שלפי האמונה מסונתזים ב"ריבוזומים".
אז מה רואים מיקרוסקופיסטים אלקטרונים אם לא ר"א? הילמן וסארטורי שיערו כי הר"א היא למעשה רק משקעים של הציטופלסמה עם חומרי הקיבוע. מכיוון שרקמה חיה מכילה 60% מים, הייבוש של דגימת הרקמה יגרום להתכווצות משמעותית גם אם המים מוחלפים לאחר מכן באלכוהולים, במגיבים (reagents) אורגניים ובמצע ההטמעה (שרף אפוקסי או שעווה). הופעת הר"א היא אפוא תוצאה של החום העז של אלומת האלקטרונים המייבשת והמשקע של רקמה המשולבת במתכת. בין אם השערה זו נכונה ובין אם לאו, המסקנה לפיה הרשתית האנדופלזמית אינה קיימת ברקמת התא החי, בעינה עומדת.
5. "ריבוזומים"
"ריבוזומים" הוא השם שניתן לגרגירים הנראים במיקרוגרפים אלקטרונים בין שכבות הר"א (ראו מיקרוגרף אלקטרונים G), ולהפרדה תת-תאית מסוימת המכילה בערך 50% של רנ"א.(מקור 16)
מקובל להאמין שהרנ"א ותפקודו הכימי בתוך התא החי קורים ב"ריבוזומים".
הילמן וסארטורי טוענים שאם הר"א הוא תוצר מלאכותי של משקעי הציטופלסמה, סביר מאוד להניח שגם הממצא של "ריבוזומים" הם[תוצאה של] משקעים.
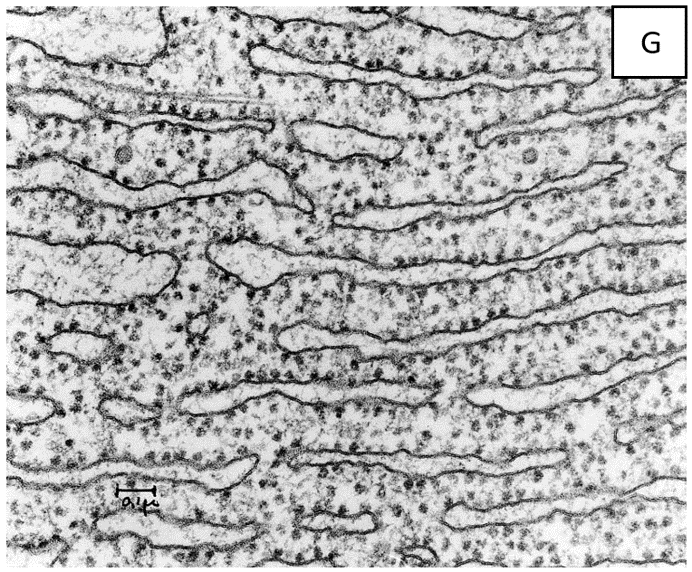
"ריבוזומים" אינם נראים במיקרוסקופ אור, והילמן וסארטורי מחזיקים בדעה כי בנוסף לבעיות שנדונו לעיל לגבי הפרדה תת-תאית, אין גם ראיה מספקת למיקום התת-תאי של פעילויות ביוכימיות. במילים אחרות, אין ראיות מספקות המוכיחות כי הרנ"א שזוהה, לאחר ההפרדה התת-תאית, מגיע באמת מה"ריבוזומים" בתא החי.
הסיבה לכך היא האמונה כי כל הממברנות "מתעגלות" למעין חלקיקים הנקראים "מיקרוזומים", ובהחלט ייתכן ש"מיקרוזומים" במקטע מסוים, שלפי האמונה הם "ריבוזומים" על גבי הר"א, יכולים למעשה להיות רסיסים של קרום התא או של כל מבנה אחר בעל קרום, שלפי האמונה נמצא בתאים. יתר על כן, ובהתחשב באמונה כי חומצת גרעין סופגת אור אולטרה סגול (UV), ולכן ניתן להשתמש באור זה כדי לזהות את חומצת גרעין,(מקור 17) הילמן מציין כי אם הר"א היתה מצופה ב- רנ"א, ניתן היה לראות זאת במיקרוסקופ אולטרה סגול כתעלות כהות המחלחלות לציטופלסמה.
במילים אחרות, המיקום בתא בו מאמינים כי ר"א ו"ריבוזומים" נמצאים, היה אמור לספוג הרבה יותר אור על סגול ולהיראות כהה יותר מסביבתו, בשל הריכוז הגבוה של רנ"א. הילמן וסארטורי לא הצליחו למצוא ראיות לכך שניסויים כאלה נעשו אי פעם.
6. "נקבוביות גרעין"
גרעיני התא (המרכז הכהה של התאים) נצפו כבר לפני 150 שנה. עם זאת, על "נקבוביות בגרעין"(מקור 18) דווח רק לאחרונה בקרומי גרעין של תאים רבים של צמחים ובעלי חיים, [והן נצפו] במיקרוסקופ אלקטרונים, אך לא במיקרוסקופ אור.
נקבובית מופיעה, בחתך רוחב של קרום הגרעין, כחוסר רציפות במראה הדו-קווי של שכבות מעטפת הגרעין (ראו מיקרוגרף אלקטרונים H). במבט משיקי הן נראות כמעגל או מתומן (ראו מיקרוגרף אלקטרונים I).
, "נקבוביות הגרעין" נחשבו בתחילה לקרעים בממברנה, אך לאחרונה יוחסו להם פרטים רבים יותר.
כיום נאמר לנו שהם חלק מ"קומפלקס נקבוביות הגרעין" המאפשר מעבר של חלבונים ורנ"א בין הגרעין לציטופלסמה, דבר בעל חשיבות מיוחדת לסינתזה של חלבונים.(מקור 19, 20)
כפי שאתם יכולים כעת כבר לדמיין, להילמן ולסארטורי יש בעיות עם תיאור הזה בדומה לאלו שהוצגו לגבי המבנים התאיים הקודמים, כלומר הראיות שהוצגו לקיומם נראות כסותרות את חוקי הלוגיקה, הגיאומטריה והביולוגיה.
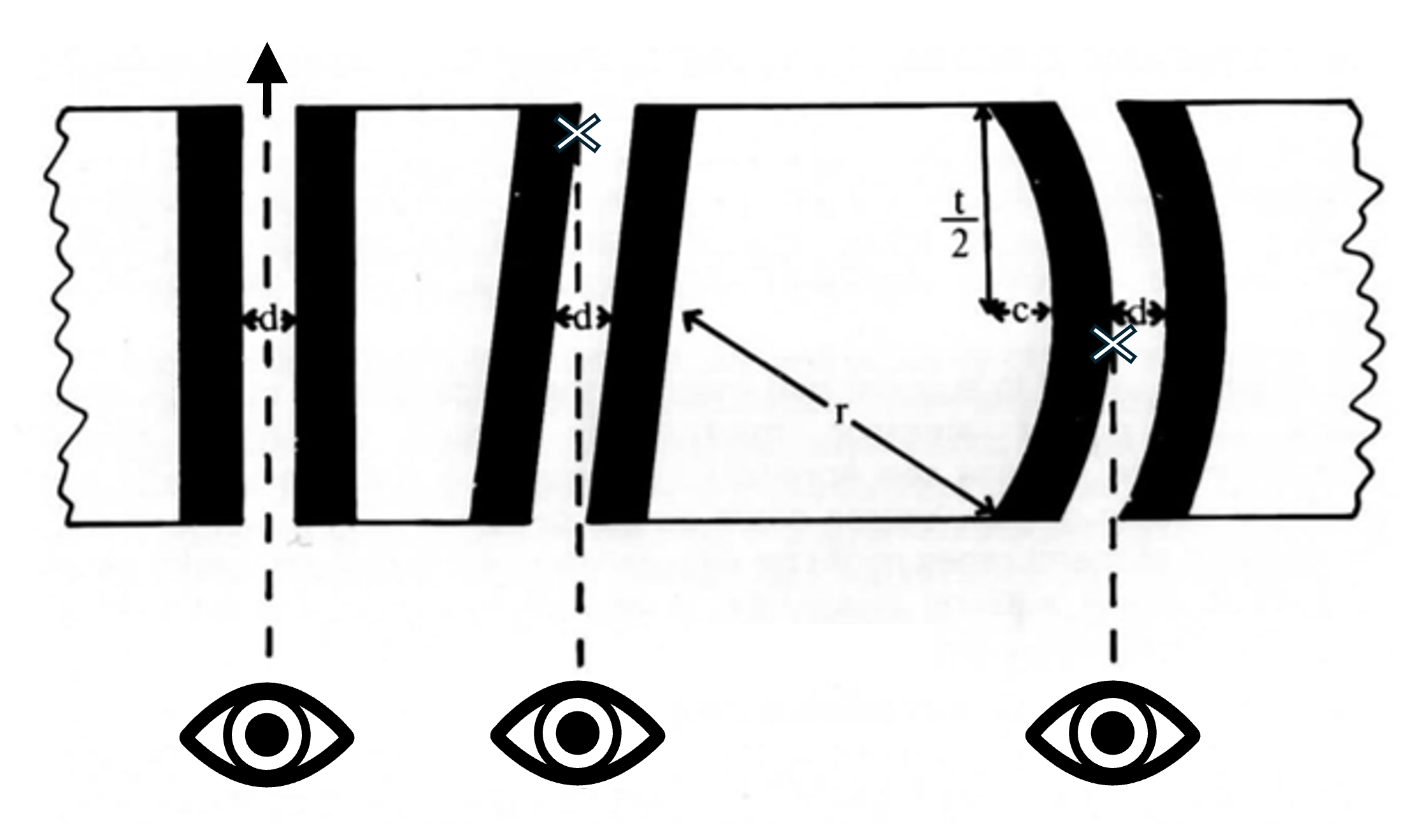
ראשית, "נקבוביות" אלו נראות תמיד רק כפערים בקרום הגרעין או כמערך של עיגולים או מתומנים. רק לעתים רחוקות הם נראים כאחת מצורות הביניים שניתן היה לצפות להן, אם בכלל. לדוגמה, ככל שה"נקבוביות" מתקרבות לקצה הגרעין הן בהכרח אמורות היו להיראות מעוכות יותר או חתוכות יותר, ולא לשמור על המראה שלהן ממש עד הקצה. ראו איור 6.
שנית, יש להבין כי ניתן יהיה לראות רווחים ב"קרום הגרעין" רק במקרים בהם קוטר "הנקבוביות" גדול מעובי החתך, ובתנאי שהחתך חותך את ה"נקבובית" משני הצדדים (פנים קדמיות ואחוריות שלה). ראו איור 7 להלן. שימו לב שהנקבובית תיראה בהכרח כאילו היא בקוטר של הפנים הקטנות ביותר שנחתכות על ידי החתך, ולכן היא לא תמיד תיראה באותו קוטר בכל חתך.
שלישית, מעגלים או מתומנים בעלי ממד דומה מופיעים בציטופלסמה, אך אלה לעולם אינם נחשבים לנקבוביות. ואכן, נראה כי הקריטריון של 'מעגל בקוטר אחיד יחסית על הגרעין' הוא הדרך היחידה לזהות "נקבוביות גרעיניות".
רביעית, גם אם, כנאמר, "נקבוביות הגרעין" מנקבות רק 3% מקרום הגרעין, קשה להבין כיצד גרעינים שומרים כנראה על צורתם הכדורית לאחר תהליכי האחדה או צנטריפוגה. שמירה על צורה כדורית מחייבת שיהיה הפרש לחצים עקבי בין החלק הפנימי והחיצוני של הגרעין – דבר שאינו אפשרי אם מעטפת הגרעין באמת מנוקבת בנקבוביות. לחילופין, צריך שיהיה מבנה פנימי [שלד] השומר את הצורה הכדורית.
חמישית, אם "נקבוביות הגרעין" מאפשרות ל-רנ"א לעבור, כיצד הן מונעות ממולקולות ויונים קטנים יותר לעבור דרכן, ו[אם אלו עוברים], אז כיצד ניתן לומר שיש הבדל פוטנציאלים (הבדל בחומציות, רמת ה-PH) על פני קרום הגרעין?(מקור 21)
במילים אחרות, כיצד נשמר ההבדל ב-PH (למעשה ההבדל בריכוז יוני מימן) בין פנים הגרעין לבין הציטופלסמה, אם הנקבוביות בגרעין מאפשרות לפילים (מולקולות mRNA) לעבור דרכם אז איך לא ליתושים (יוני מימן)?
הילמן וסארטורי טוענים כי סביר הרבה יותר ש"נקבוביות הגרעין" הן קרעים בקרום הגרעין הנוצרים בתהליכי הקיבוע, היבוש, ההקפאה או השימוש בקרן האלקטרונים.
7. מודל מדויק של התא החי
חשוב לשים לב שגם "ליזוזומים" ו-"מנגנון גולג'י", ששניהם נחשבים כנמצאים בציטופלסמה, וגם ה"כריסטות" [מבנה דמוי מחיצה מפותלת] שאמורות להיות ממוקמות בתוך המיטוכונדריה, הם כולם אברונים שנאמר כי נמצאים בתא החי לאחר שנתגלו רק עם מיקרוסקופ אלקטרונים. הם אינם נדונים כאן מכיוון שהם אינם רלוונטיים לענין המאמר. עם זאת, על הקורא לדעת שהילמן וסארטורי הדגימו כי גם אברונים אלה אינם אלא תופעות מלאכותיות [של תהליך הצפייה], וזאת מסיבות דומות לאלו שפורטו לעיל, דהיינו, נראה שהם סותרים את כללי הלוגיקה, הגיאומטריה והביולוגיה.(מקור 22)
בסיכומו של דבר, הילמן וסארטורי מראים כי הראיות לכך שקרום התא, המיטוכונדריה, הציטופלסמה והגרעין אכן קיימים בתא החי, הן מעבר לספק סביר. לא כך לגבי שאר האברונים שמקובל לחשוב עליהם כחלק מהתא החי. נראה כי אלו רחוקים מלהיות ודאיים.
אסכם שוב עבור אלה שמתקשים להתרכז. למעט הגרעין והמיטוכונדריה, כל שאר האברונים הנחשבים כנמצאים בתאים של צמחים ובעלי חיים, הן תופעות מלאכותיות המיוצרות על ידי מיקרוסקופ האלקטרונים, והם אינם קיימים מחוץ למיקרוגרפים אלקטרונים. מודל התא הכללי מלפני 1940, הוא המודל היחיד של תא חי [שאפשר להתיחס אליו] כמדויק.
8. סינתזת חלבונים, הדוֹגמה המרכזית של גנטיקה ודנ"א
כפי שנרמז בדיון לעיל בעבודתו של הילמן, התפקיד העיקרית המיוחס לאברוני התא ש"התגלו" במיקרוסקופ אלקטרונים מתייחס לייצור חלבונים. כיצד סינתזת חלבונים הפכה לשם-נרדף של ביטוי גֶנים והדוגמה המרכזית של הגנטיקה אסביר בפירוט רב יותר בהמשך, אך תחילה יש צורך לבחון בקצרה כיצד ביולוגיים מולקולריים קבעו כי סינתזת חלבונים אכן מתרחשת בתאים. ראוי גם לציין כי לפי האמונה המקובלת, התא החי מכיל בעיקר ממים, חלבונים ואשלגן, והוא צף בתוך סביבה של מים ונתרן.
א. סינתזת חלבון ללא תאים (CFPS)
המנגנונים האמורים להיות אחראים לסינתזה של חלבונים התגלו על ידי ביוכימאים וביולוגים מולקולריים בשנות ה- 1950-60. שניים מההיבטים החשובים ביותר היו גילוי הרנ"א (tRNA ו-mRNA) והתובנה לפיה הרנ"א נמצא ברשתית האנדופלזמית (ר"א) וב"ריבוזומים" של כל התאים.
תגליות מכריעות אלה הן שהובילו ביוכימאים וביולוגים מולקולריים ליכולת לסנתז חלבונים במבחנות על ידי הדגרה של הפרדות "ריבוזומליות" (רנ"א), הידועות גם בשם "מיקרוזומים", בתוך תערובות מסוימות של מצעים – חומצות אמינו (אבני בניין של חלבונים), מלחים (נתרן ואשלגן כדי לדמות סביבה של תאים חיים) ואדנוזין טריפוספט (ATP).
ATP היא מולקולה גדולה אשר אמורה להיווצר על ידי המיטוכונדריה בתוך התאים מתוך גלוקוז שמקורו במזון שאנו צורכים (תהליך הידוע בשם גליקוליזה). נאמר עליה שהיא מתפקדת כמולקולה האוצרת אנרגיה רבה שמשמשת לפעילות בתאים. כאשר הכימיים הקשרים של מולקולת ה-ATP נשברים באמצעות הידרוליזה חומצית בתוך "ליזוזומים", משתחררת אנרגיה אשר הנרתמת על ידי "ריבוזומים" ליֶצור (סינתזה) של חלבונים. דבר זה מביא לאמונה כי ATP הוא מקור האנרגיה המניע סינתזת חלבונים.(מקור 23)
בגדול, המנגנונים של סינתזת חלבונים פוענחו בניסויים שהחלו בנוזל אחוד [הומוגנט] של התא שיכול היה לסנתז חלבונים. ההפרדה תת-תאית של תמיסת תא אחודה אפשרה להוסיף לתערובת המצע המודגרת סוגי חלקיקים מופרדים שונים בנפרד זה מזה או למנוע אותם, כדי לקבוע את תפקידם המדויק בתהליך ולזהות אילו שילובים מובילים לכך שחומצות האמינו במבחנה מסונתזות לכדי חלבונים.
הליך זה ידוע בשם "סינתזות חלבון ללא תאים" (CFPS) ומהווה את הבסיס לביולוגיה סינתטית. החלקיק התת-תאי הדרוש לסינתזה של חלבונים במבחנות זוהה מאוחר יותר באמצעות מיקרוסקופ אלקטרונים כנובע מהר"א ומה"ריבוזומים".
השאלה האם תהליך CFPS מדגים מעבר לכל ספק שתאים מסנתזים חלבונים, היא נושא למאמר אחר. אך בהתחשב בכך שהניסויים העומדים בבסיסם היו אפשריים רק באמצעות הליכי הפרדה תת-תאית, שימוש במיקרוסקופ אלקטרונים וטיפולים כימיים – שלגבי אף אחד מהם אין הוכחה הנשענת על ניסויי ביקורת, המראה כי ההליכים עצמם אינם משפיעים על תוצאות הניסויים (כפי שנדון לעיל במאמר זה) – יהיה זה אך הוגן לומר שתגליות אלה נשענות על יסודות רעועים.
למרות זאת, ניתן להם לעת עתה ליהנות מהספק ונקבל, לצורך מאמר זה, שתאים אכן מסנתזים חלבונים. מה שלא ניתן לקבל, עם זאת, זה שהחלבונים אכן מיוצרים על ידי הר"א וה"ריבוזומים" בתאים. וכך אכן מעידה עבודתו של הילמן.
למי שתוהה, חלבונים אמורים לבצע מגוון רחב של תפקידים חיוניים באורגניזמים חיים. הם פועלים כאנזימים, הורמונים, נוגדנים, רכיבי מבנה ומולקולות הסעה. סינתזת חלבונים מאפשרת לתאים לייצר רכיבים נחוצים אלה בתגובה לצרכים משתנים, כגון גדילה, תיקון וויסות תהליכים פיזיולוגיים. בעיקרון, חלבונים קובעים כיצד הגוף שלך בנוי וכיצד הוא מתפקד.
ב. ביטוי הגֶנים והדוֹגמה המרכזית
ביטוי-גֶנים מהווה את הדוֹגמה-המרכזית של ביולוגיה מולקולרית וגנטיקה, ומאז שפרנסיס קריק הציע אותו לראשונה לפני למעלה מחמישים שנה, מודל הדוֹגמה-המרכזית השתלט על המחקר בתחומים אלה. במילים פשוטות, הדוֹגמה-המרכזית היא סיפור המקור עבור רנ"א, שבלעדיו סינתזת חלבונים (נטולת תאים) לא הייתה יכולה להתקיים.
הדוגמה-המרכזית קובעת שהדנ"א הוא סטטי, הוא אינו משתנה מעצמו מרגע שנוצר; מדנ"א נוצר רנ"א ('שעתוק'); רנ"א מתורגם לחלבונים על ידי מכונות ייצור החלבונים בתאים ('תרגום'). תהליך זה יכול לזרום רק בכיוון אחד ולא ניתן שחלבונים יומרו בחזרה להיות חומר גנטי. לבסוף, מאחר שהדנ"א מורכב תמיד מאותם ארבעה נוקלאוטידים בכל יצור חי, הקוד הגנטי הוא אוניברסלי, מה שאומר שגֶן צריך להיות מסוגל לייצר את החלבון המסוים שלו בכל מקום שבו הוא מוצא את עצמו, אפילו במין אחר.
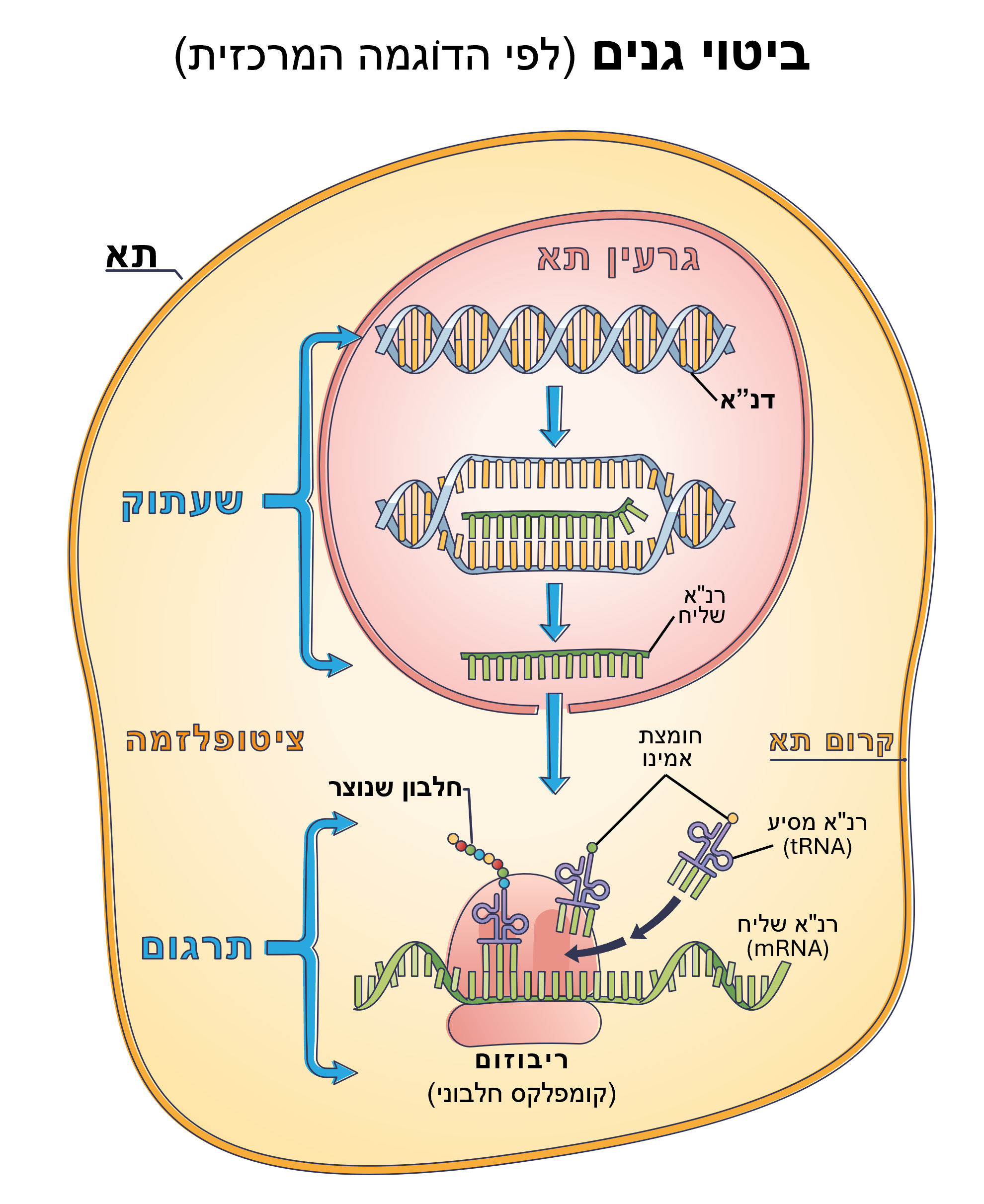
סינתזת חלבונים, המבוצעת על ידי הר"א וה"ריבוזומים", נחשבת על כן כנמצאת תחת שליטה בלעדית של הדנ"א שלנו ושל הגֶנים שמהם הוא מורכב. הסיבה לכך היא האמונה שרצף הנוקלאוטידים בגֶן "הוא קוד פשוט (מתכון) עבור רצף חומצות אמינו של חלבון מסוים".(מקור 24)
הנה הסבר קצר של תהליך ביטוי הגֶנים: אנזים (סוג של חלבון), המכונה רנ"א פולימראז, אחראי על שעתוק של גן (מקטע של דנ"א) לתוך מה שנקרא רנ"א-שליח (mRNA).
לאחר יצירתו, עוזב רנ"א-שליח זה את הגרעין דרך "נקבוביות הגרעין" אל תוך הציטופלסמה, וצף בתא בדרכו למנגנון ייצור החלבונים של התא. כפי שאתם כבר יודעים, "הרשתית האנדופלסמית" ובמיוחד ה"ריבוזומים" הם האברון שאמור להיות אחראי על סינתזת חלבונים ולכן גם על תרגום הרנ"א-שליח והרכבת חומצות אמינו לשרשראות חלבונים (פפטידים) בהתאם למתכון שהגיע עם מולקולת הרנ"א-שליח.
מולקולה המכונה רנ"א-מסיע (tRNA) נחשבת כמחוברת לחומצות האמינו ומתפקדת כמתרגם או מתאם עבור "ריבוזומים". זאת מכיוון שה"ריבוזומים" אמורים לקרוא את קוד הרנ"א-שליח, שלושה זוגות של נוקלאוטידים (קודונים) בכל פעם, ואז להתאים אותם לקודונים המשלימים (אנטיקודונים) שנמצאים על מולקולת הרנ"א-מסיע.(מקור 25)
חשוב לציין שאף אדם לא צפה מעולם בדוֹגמה-המרכזית בפעולה בתוך תא חי כלשהו. תחת מיקרוסקופ אור וברקמה לא מקובעת (רקמה חיה), לא ניתן לראות אף לא אחד מהמבנים והמולקולות שנטען כי הם מעורבים [בתהליך זה]. המבנים ניתנים לצפייה רק במיקרוסקופ אלקטרונים ואת המולקולות ניתן למצוא רק לאחר ביצוע הפרדה תת-תאית.
הדוֹגמה-המרכזית היא אפוא מודל תיאורטי שהוצע לאחר שנעשו תצפיות על א) רקמות שעברו פרוצדורות וטיפולים כימיים שונים, וב') תמונות סטטיות שהתקבלו ממיקרוסקופ אלקטרונים. יתר על כן, קשה מאוד להאמין שתהליך הסינתזה אמור להתרחש בדיוק באותו אופן במבחנה לאחר הפרדה תת-תאית.
ג. דנ"א
דנ"א, שממנו לכאורה מגיע הרנ"א, אמור להיות מולקולה ליניארית ארוכה מאוד המעוצבת כסליל כפול המלופף בצפיפות בתוך גרעין התא.
האמונה היא כי דנ"א מורכב מארבעה סוגים שונים של נוקלאוטידים, אשר בכל גן הם המחוברים יחד בסדר ליניארי או רצף מסוים. ארבעת סוגי הנוקלאוטידים יכולים להיות מסודרים ברצפים אפשריים רבים, והסידור שלהם בהרכב של גן מסוים מייצג את "המידע הגנטי"(מקור 24)
האמונה היא שכל תא אנושי מכיל בערך 2 מטרים של דנ"א האמור להיות ארוז בגרעין שגודלו 6 ננומטר. זה דומה לאריזה של 40 ק"מ של חוט דק במיוחד לתוך כדור טניס.
דבר זה אפשרי לכאורה, מכיוון שארבעה מחרוזות דנ"א דו-גדיליות נכרכות ומקופלות בחוזקה סביב מולקולת היסטון (סוג של חלבון) לחבילות הידועות כנוקלאוזום. האמונה מציעה כי נוקלאוזומים נדחסים עוד יותר למבנה ספירלי המכונה כרומטינים, אשר לבסוף נדחסים עוד יותר לכרומוזומים.
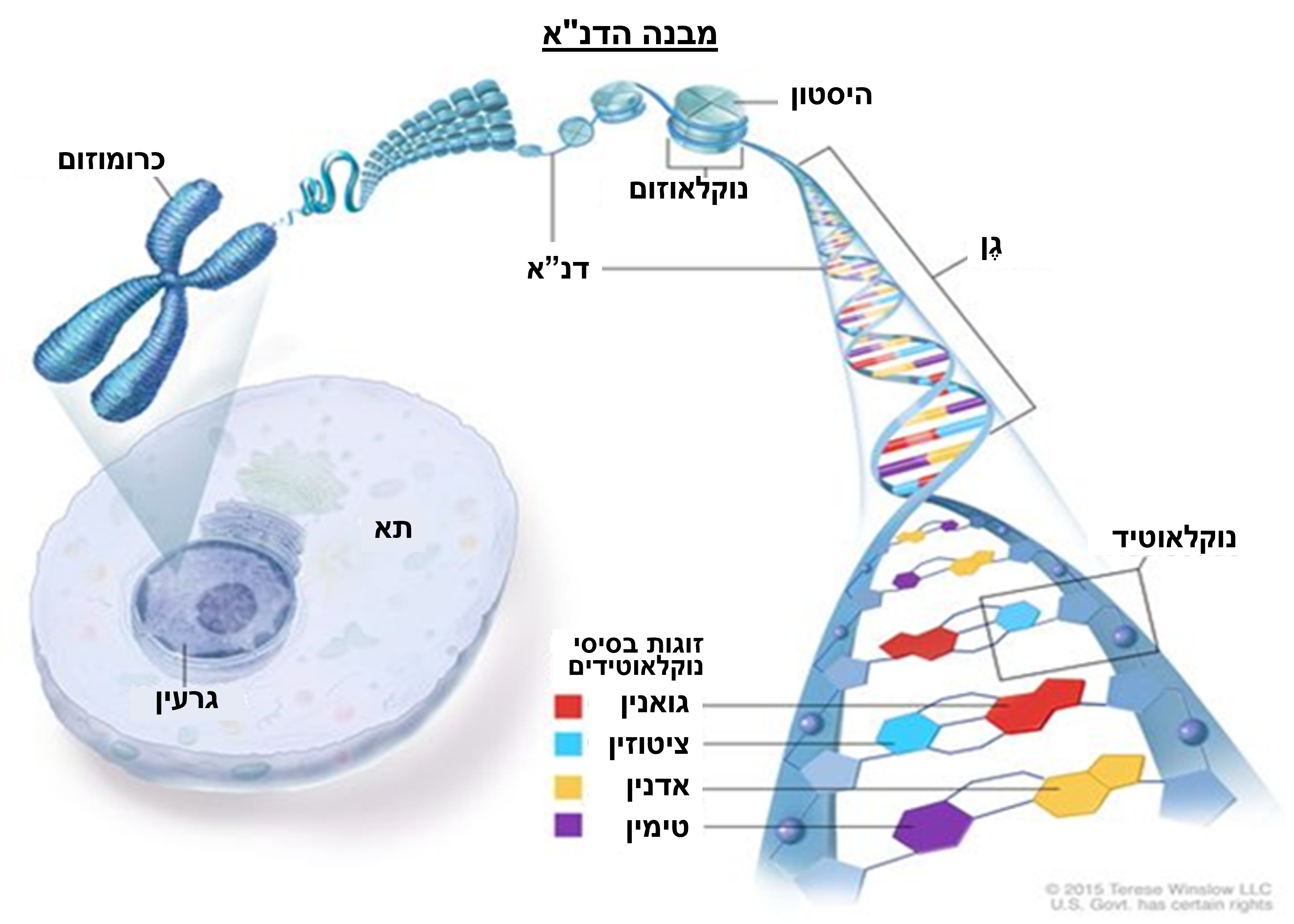
הטענה מציעה כי לבני אדם יש 46 כרומוזומים המתקיימים בזוגות, אחד מהאבא שלנו ואחד מהאמא, וכי בתוך גרעיני התא שלנו ישנם 23 כרומוזומים זוגיים במבנה הדומה בערך למוט קווי. סך הרצף הכולל של מחרוזות דנ"א אלה בתוך הכרומוזומים מוכר בשם הגנום האנושי (מעין תבנית).(מקור 24)
למרבה הפלא, למרות היותם מקופלים היטב ועטופים ודחוסים למבנים מורכבים, אין ככל הנראה כל בעיה כאשר יש צורך להעתיק את הגדילים ולשעתק אותם ב-mRNA.(מקור 26)
חשוב לציין כי ביולוגים מולקולריים מרגישים מוסמכים להצהיר הצהרות אלה לגבי הרכב הדנ"א ומבנהו, מכיוון שלטענתם הם בודדו את הדנ"א שבתאים. במובנים רבים, טענות אלה דומות לאלה המושמעות לגבי המבנה הפנימי של תאים, בכך שהן אינן לוקחות בחשבון את ההשפעה של הליכי הניסויים שהובילו לתגלית, על הדברים הממשיים שלטענתם נתגלו.
אינני יכולה לבחון ביקורתית את הטענות הללו בצורה טובה יותר מכפי שעשתה כבר TAM במאמרה בנושא,(מקור 27) אני חושבת שמאמרה של TAM הוא קריאת חובה לכל מי שרוצה לדעת על דנ"א.
השורה התחתונה היא, שלא ניתן לטעון ביושר כי מה שבודָד על ידי ביולוגים מולקולריים הוא דבר שונה מאותם תוצרי הלוואי שנותרים בהמשך לערבוב דגימה ביולוגית עם כימיקלים וחימום שלה. הדבר נכון במיוחד לאור העובדה שלא בוצעו כל ניסויי ביקורת. אם עצם בידוד הדנ"א הוא מפוקפק, מבנה המולקולה הוא, במשתמע, מפוקפק במידה דומה.
ד. הדוגמה המרכזית ותורשה
חיוני להבין שהדוֹגמה-המרכזית מצמצמת את היווצרות החיים ואת התורשה לכדי תגובות כימיות בממדים מולקולריים, זאת מאחר והם הצביעו על דנ"א כעל המתווך המולקולרי של התורשה.
מה שהוביל למסקנה זו היא העובדה שכאשר מתבוננים בתאים המתחלקים (מיטוזה) תחת מיקרוסקופ אור, ניתן לראות חלקיקים חוטיים בגרעין התא מתחלקים ואז נפרדים כאשר חצי אחד נשאר בתא המקורי והשני נע לתוך הגרעין של תא הבת שזה עתה נוצר. בהמשך הוצע כי חלקיקים חוטיים אלה הם כרומוזומים והם הסיבה לכך שכל התאים העתידיים יצרו חלבונים זהים.
שימו לב, הילמן ציין שניתן לצפות בתופעה זו רק בתאי עובּר וכי לא בכל התאים החיים ניתן לראות כרומוזומים.(מקור 28) תאי עובר נמצאים רק בעובּרים או בתינוקות שטרם נולדו.
האמונה היא, על כן, כי דנ"א (ארוז בכרומוזומים) הוא ספר המתכונים של איך להרכיב אדם מסוים. המתכונים בספר (הגֶנים שלנו) הם שילוב של מתכונים שנלקחו ככל הנראה מספרי המתכונים של אבא שלנו או מזה של אמא שלנו. במילים אחרות, התאים שלנו יכולים לייצר חלבונים רק בהתאם למתכונים שהתקבלו מאחד ההורים שלנו.
כלומר, אם להורים שלנו, או לכל אחד אחר בשושלת שלנו, יש מתכונים פגומים או ייחודיים, יש סיכוי שהמתכונים האלה יועברו אלינו והתאים שלנו ייצרו חלבונים פגומים או ייחודיים, שתהיה להם, בתורם, השפעה מיוחדת מסויימת על המבנה או התפקוד של הגוף שלנו.
9. הערות סיכום
כאשר פורסים את עבודתו של הילמן הנוגעת לתא החי, על התיאוריה של הדוגמה-המרכזית והסינתזה של חלבונים כפי שהיא מקובלת כיום, מתברר כי מודלים אלה אינם יכולים לעבוד כפי שמקובל להאמין. המבנים התאיים שאמורים להיות מעורבים בתהליך ואשר נותנים אמינות לתיאוריה, פשוט לא קיימים.
כאשר הילמן ניסה להצביע על כך בפני הקהילה המדעית, הוא נתקל בעיקר בעוינות, וכל כתבי העת המדעיים "המכובדים" והידועים ביותר סירבו לפרסם את מאמריו בנושא. כתוצאה מכך, הילמן ועבודתו זכו להתעלמות מוחלטת ונקברו תחת הררי "מדע" לא מדויק.
א. סוגיות נוספות עם סינתזת חלבונים והדוגמה המרכזית
בנוסף לעובדה ש"הרשתית האנדופלסמית" וה"ריבוזומים" אינם קיימים בתא החי, חשוב להיות מודעים לכמה סוגיות מכריעות נוספות שהילמן הצביע עליהן במודל המקובל כיום של סינתזת חלבונים.
ראשית, איפה בדיוק מתרחשת סינתזת חלבונים בתאים, שאין בהם "ריבוזומים"?(מקור 29)
שנית, מהן הראיות לטענה שכל תא של צמח או של בעל חיים מסוים, מכיל בדיוק את אותה כמות של דנ"א, במיוחד לאור כך שיש שונות עצומה בגודלם של גרעיני תאים?(מקור 30)
אם ניתן להראות כי לתאים מחלקים שונים של גופנו יש כמויות שונות של דנ"א – כפי שמוצג במאמרים אלה הדנים במחקרים אחרונים בנושא זה(מקור 31) – הרי שהדוֹגמה-המרכזית (גֶן אחד שווה חלבון אחד) ו"שותפתה לדבר עבירה", סינתזת חלבונים באמצעות "ריבוזומים", אינן יכולות עוד להיחשב כעובדות שאינן ניתנות להפרכה?
שלישית, עד למועד בו יבוצעו ניסויי ביקורת מתאימים להליך "הפרדה תת-תאית", לא ניתן יהיה לומר באופן בלתי ניתן להפרכה כי הדנ"א ממוקם בגרעיני התאים.(מקור 32) שימו לב עוד לכך שדנ"א ורנ"א הופקו עד כה אך ורק מרקמות מתות.
את הנקודות הבאות הילמן לא העלה , אך הן בכל זאת רלוונטיות לדיון זה:
רביעית, אם הדוגמה-המרכזית נכונה, המסקנה היא שבכל יצור חי צריכה להיות התאמה של אחד לאחד בין מספר הגֶנים הכולל לבין המספר הכולל של החלבונים. כל מערך הגֶנים האנושיים, כלומר הגנום, חייב אפוא לייצג את כל התורשה, או התבנית, של האדם, אשר היא המבדילה בין אדם לזבוב.
בשנות ה-90 של המאה ה-20 יצא פרויקט הגנום האנושי לזהות ולמנות את כל הגנום האנושי על ידי חישוב הרצף של 3 מיליארד הנוקלאוטידים בדנ"א האנושי. אולם, להפתעתם, ממצאיהם סתרו את הדוֹגמה-המרכזית לפיה גן אחד שווה לחלבון אחד. במקום למצוא את 100,000 הגֶנים או יותר המבוססים על המספר המשוער של חלבונים אנושיים, הם הצליחו למצוא רק כ -30,000 גֶנים.
למען הפרספקטיבה, זה הופך את בני האדם למצוידים-גנטית כמו צמח עשב החרדל, שיש לו 26,000 גֶנים, ומצוידים-גנטית רק פי שניים מזבוב פירות.(מקור 24) ככאלה, ה"גֶנים" שלנו אינם יכולים להסביר את ההבדל התורשתי העצום בין עשב לבין אדם. הדבר בולט אם לוקחים בחשבון את העובדה שלבני אדם, הנחשבים למורכבים ביותר מבין יצורי כדור הארץ, יש פחות גֶנים מאשר לבעלי חיים וצמחים.(מקור 33)
חמישית, נראה כי הטענה שהדנ"א הוא סטטי הופרכה לפני עשרות שנים על ידי ברברה מקלינטוק, כאשר תיארה כיצד רכיבים "חוץ-גניים", המכונים טרנספּוזונים, יכולים לשנות את תפקודם של גֶנים סמוכים.(מקור 34) כלומר, המנגנון השולט בגֶנים יכול להיות בלתי תלוי בגֶנים עצמם. למרבה האירוניה, מסקנותיה של מקלינטוק לגבי חוסר יציבות הדנ"א פורסמו באותה שנה [1953] שבה פורסמו דוחות המתארים את מבנה הסליל הכפול של הדנ"א מאת פרנסיס קריק וג'יימס ווטסון.
שישית, ממחקרים אחרונים נראה כי ביוכימאים ממשיכים למצוא זוגות חדשים של בסיסי נוקלאוטידים, ואכן נטען כי למעשה עשויים להיות שבעה או שמונה זוגות של בסיסים אלו.(מקור 35, 36) דבר זה קורא תיגר מוחלט על הבנתנו את מבנה הדנ"א ואת תפקידו בדוֹגמה-המרכזית של הגנטיקה.
ב. איפה זה משאיר אותנו?
תקוותי כי הדיון לעיל על [השיטות המשמשות] במדע המקובל מבהיר כי:
- תאים חיים אינם מופיעים בטבע כפי שהם נראים במיקרוגרפים אלקטרונים;
- למעט המיטוכונדריה והגרעין, אין אברונים [אחרים] בתוך התא החי;
- הר"א וה"ריבוזומים" אינם מייצרים חלבונים בהתאם למולקולות mRNA המעוצבות ע"פ הגֶנים והדנ"א שלנו;
- הגֶנים שלנו אינם מתכונים לחלבון יחיד;
- את הדוֹגמה-המרכזית של הגנטיקה צריך לגרוט במגרש הגרוטאות; ו..
- כל הרעיון של הדנ"א צריך לעבור הערכה מחדש.
ג. מהן כמה מההשלכות היומיומיות של תגליות אלו?
ראשית, עכשיו צריך כבר להיות ברור לחלוטין שהגֶנים אינם אחראים לתכונות התורשתיות שלנו. לא הוכח כי דנ"א שולט בסינתזת חלבונים, ובנוסף, לא הוכח שהדנ"א הוא סטטי או זהה ועקבי בכל תא ותא.
פירוש הדבר הוא לא רק שהגֶנים אינם קובעים את צבע השיער והעיניים שלנו, וכדומה, אלא גם שהם לא יכולים להיות הגורם האחראי לרגישות שלנו ל"מחלות" מסוימות – כמו למשל מחלות אוטואימוניות ומחלות פסיכיאטריות.
שנית, זה גם אומר ששיבוט הוא שטות מוחלטת, כי אין גנום אנושי שאפשר לשכפל ולהרכיב, וגם אם היה, אין לנו שום מושג איך חלבונים נוצרים. שימו לב: אני לא טוענת שתורשה של תכונות אינה מתרחשת, אלא פשוט מציינת שאין לנו מושג איך זה קורה.
שלישית, בראותינו כי וירוסים הם עיקר הטירוף של הזמן האחרון, הבה נמשיך ונטפל בהם כעת. וירוסים אמורים להיות פיסות של חומר גנטי עטופות במעטפת חלבונית, הפועלים כטפילים תוך-תאיים המשתלטים על מנגנוני ומסלולי יצור החלבונים של התא (הריבוזומים והרשתית האנדופלסמית) כדי לשכפל את עצמם.
בעיקרון, בין אם נאמר שהם וירוסי רנ"א או דנ"א, התהליך זהה. הווירוס עושה את דרכו לתוך התא ומחדיר את החומר הגנטי שלו למנגנון ייצור החלבונים של התא, כך שבמקום לייצר את החלבונים הדרושים לאורגניזם החי, התא מייצר כעת את החלבונים הוויראליים הדרושים לשכפול הווירוס. ובכן, אם מנגנון יצור החלבונים אינו קיים בתא, כיצד בדיוק יכול שכפול להתרחש?
אם משלבים את זה עם העובדה ש[שום] ווירוס מעולם לא בודד(מקור 37, 38) ועם מחקרי הדבקה [במחלות] הרבים שהתרחשו [וכשלו] במהלך 150 השנים האחרונות (מקור 39) , וירוסים הם כנראה לא יותר מאשר סיפור אגדה של וירולוגים, וכל זריקות ה"חיסונים" אינם אלא בדיחה חולנית.
עוד סיבה לפסאודו-מדע הזה היא שהוא מהווה את הבסיס התיאורטי מאחורי מיליארדי הדולרים של תעשיית גידולי מזון מהונדסים גנטית (GMO). כאשר הם מהנדסים גנטית צמחים, ביולוגים טוענים שהם מכניסים גֶנים מועילים לדנ"א של צמח ובכך מספקים לצמח את המתכון או הקוד הדרושים לייצור חלבונים, שיאפשרו להם להיות חסינים מפני ההשפעות של קוטלי חרקים וקוטלי עשבים מסוימים. עם זאת, ברור מהדיון לעיל כי, א) לתאים אין מנגנון לייצור חלבון, ב) גֶנים לא הוכחו כשולטים בסינתזת חלבונים ג) גם אם קיימים גֶנים כמתואר, סביר יותר שהם אחראים ליצירת סוגים רבים ושונים של חלבונים ולא רק אחד.
ריפוי גנטי הוא פחות או יותר רק ניסיון לשנות גנטית בני אדם במקום גידולי מזון, וככזה, הוא נתקל באותן שלוש בעיות שפורטו לעיל עם הנדסה גנטית. לדוגמה, מה אם אותם חיסונים שאמורים להכיל mRNA ויכולים להביא לריפוי גנטי?
נאמר כי מטרת ה-mRNA בחיסונים מסוימים היא להורות לתאים שלנו לייצר חלבון מסוים, חלבון הסְפַּייק הוויראלי, שיגרום לגופנו בתורו לייצר את הנוגדנים המתאימים שיגנו על הגוף מפני וירוס מסוים. ריפוי גנטי, לעומת זאת, כרוך בביצוע שינויים מכוונים בדנ"א של האדם על ידי החדרת גן המאפשר לתאים לייצר חלבון ספציפי המרפא או מקל על מצב גנטי.
ריפוי גנטי וחיסוני mRNA מתכנסים יחד בגלל חשש שאם ה-mRNA מהחיסונים לא יומר לחלבוני ספייק, הוא עלול לחדור לגרעין התא דרך "נקבוביות הגרעין" ולהתחבר לדנ"א המהודק בצפיפות שלנו, ובכך לשנות לצמיתות את הדנ"א שלנו ואת כל תאי הבת שירשו אותו משם ואילך. כאן שוב, אם אין מפעלי חלבונים בתאים ואם "נקבוביות הגרעין" אינן קיימות, איך כל זה אמור להתרחש?
לבסוף, יש לומר שכל מחקר מדעי הלוקח את מבנה התאים, את הדנ"א ואת תהליך סינתזת החלבונים, כמובנים מאליהם – כמו לדוגמה וירולוגיה וגנטיקה, אינו אלא בזבוז זמן משווע. דבר זה כולל את כל הזמן המושקע בלימוד נושאים אלה באוניברסיטה. שני ענפים אלה של מדעי רפואה אינם אלא "ביוב מוחות" מאחר והם ממש מנקזים אנשים מהתמקדות בנושאים בעלי מהות.
ד. סיכום
אם תלמדו רק דבר אחד בסיום קריאת מאמר זה, מי יתן ותהיה זו העובדה שהרוב המכריע של העבודה והמחקר של ביוכימאים, ביולוגים מולקולריים ומיקרוסקופיסטים-אלקטרונים (כלומר, אותם אנשים שמדברים על דנ"א, רנ"א, גֶנים ומולקולות) שבוצעה במהלך 100 השנים האחרונות אינו מדע בדוק ואינו ראוי לאמון. בין אם הם רימו את עצמם או פשוט אינם ישרים אינטלקטואלית, זה אינו ידע שבריאותנו וחיי היומיום שלנו צריכים להתבסס עליו.
עבודתם של מדענים אלה לא עשתה דבר מלבד לעורר פחד ולהעביר סמכות אינטלקטואלית לאנשים שאינם ראויים, והיא צריכה להיפסק. האנושות נעשית יותר ויותר משועבדת לטיפשות ולחומרנות שלה עצמה.
הדרך היחידה לצאת מזה היא לקחת בחזרה את הידע מידיהם של כמה מומחים ולהחזיר אותו לידי הרוב. הדרך לעשות זאת היא להפוך את הדברים למובנים עבור הרוב ולא ליותר מסובכים ובלתי מובנים. אני מקווה שהמאמר הזה הוא צעד בכיוון הזה.
רשימת מקורות
מקור 1 – הספד להרולד הילמן. [אנגלית]
מקור 2 – המחיר של יושרה אינטלקטואלית – שאל את הנוירוביולוג – הרולד הילמן. [אנגלית]
מקור 3 – הילמן, הרולד, "ודאות וחוסר ודאות בטכניקות ביוכימיות" [אנגלית] – Hillman, H. (1972). Certainty and Uncertainty in Biochemical Techniques. Surrey University Press, Henley on Thames. [אנגלית]
מקור 4 – פודקסט של תום קואן על הרולד הילמן ו- 47 השאלות (התחלה Tom Cowan podcast on Harold Hillman’s 47 Questions (start at 03:26. [אנגלית]
מקור 5 – ספר: "התא החי" מאת ה. הילמן, פ. סארטורי פרק Hillman, H. and Sartory, P. (1980). The Living Cell. Packard Publishing Company, Chichester (“Living Cell”) – 1 . [אנגלית]
מקור 6 – ספר: "התא החי" דפים 35 – 43
מקור 7 – "מבנה העל של תאי חיות", עמודים The Ultrastructure of the Animal Cell L. T. Threadgold. Elsevier Science & Technology, 1976 pgs 64 – 75 להסבר על מבנה הכריך של ממברנת התא,
וגם Cell Membranes – The Cell – NCBI Bookshelf לביוכימיה של ממברנת התא. [אנגלית]
מקור 8 – "ביולוגית תא מבוססת ראיות, עם כמה השלכות למחקר הרפואי" – Hillman, H. (2008). Evidence-based Cell Biology, with some Implications for Clinical Research. [אנגלית]
מקור 9 – סרטון הסבר על הפרדה בצינטרפוגה [אנגלית]
מקור 10 – הפרדה תת-תאית . [אנגלית]
מקור 11 – רשימת 23 השערות בבסיס הפרדה תת-תאית מתוך "ודאות וחוסר ודאות בטכניקות ביוכימיות" – Certainty and Uncertainty in Biochemical Techniques, Harold Hillman Surrey University Press, 1972.. [אנגלית]
מקור 12– שאלה 12 מתוך הרולד הילמן: 47 שאלות ב[מחקר] בביולוגיה שנותרו ללא מענה. [אנגלית]
מקור 13 – ספר: "התא החי" דפים 43 – 60 – Hillman, H. and Sartory, P. (1980). The Living Cell. Packard Publishing Company, Chichester (“Living Cell”) . [אנגלית]
מקור 14 – "רשתית אנדופלזמית" מתוך "ביולוגיה מולקולרית של התא" – Molecular Biology of the Cell. 4th edition. [אנגלית]
מקור 15 – "מבנה רשתית אנדופלזמית וקשריה עם איברונים אחרים" – Amber R. English, 2013. Endoplasmic Reticulum Structure and Interconnections with Other Organelles.[אנגלית]
מקור 16 – ספר: "התא החי" דפים 60 – 61 – Hillman, H. and Sartory, P. (1980). The Living Cell. Packard Publishing Company, Chichester (“Living Cell”) . [אנגלית]
מקור 17 – "מיפוי מסיבי של חומצת גרעין וחלבונים עם מיקרוסקופית על-סגול עמוקה בתא-חי" – Benjamin J Zeskind, 2007. Nucleic acid and protein mass mapping by live-cell deep-ultraviolet microscopy [אנגלית]
מקור 18 – ספר: "התא החי" דפים 67 -75 [אנגלית]
מקור 19 – מעבר מולקולות בין גרעין התא לציטוזול – Molecular Biology of the Cell. 4th edition. [אנגלית]
מקור 20 – המעטפת התאית ותנועה בין הגרעין לציטופלזמה – The Cell: A Molecular Approach. 2nd edition. [אנגלית]
מקור 21 – שאלה 32 מתוך הרולד הילמן: 47 שאלות ב[מחקר] בביולוגיה שנותרו ללא מענה. [אנגלית]
מקור 22 – ספר: "התא החי": ליזוזומים (דפים 66-67), גופיפי גולג'י (דפים 61 – 66), כריסטות (דף 61). [אנגלית]
מקור 23 – כיצד תאים משיגים אנרגיה. [אנגלית]
מקור 24 – חשיפת מיתוס הדנ"א: היסודות המופרכים של ההנדסה הגנטית. [אנגלית]
מקור 25 – מרנ"א לחלבון. [אנגלית]
מקור 26 – דנ"א כרומוזומלי. [אנגלית]
מקור 27 – גילוי דנ"א מיצויו והמבנה שלו. בדיקה ביקורתית [עברית]
מקור 28 – עידן חומצות האמינו והרופא ה(לא)נורמלי החדש. [אנגלית]
מקור 29 – שאלה 7 מתוך הרולד הילמן: 47 שאלות ב[מחקר] בביולוגיה שנותרו ללא מענה. [אנגלית]
מקור 30 – שאלה 33 מתוך הרולד הילמן: 47 שאלות ב[מחקר] בביולוגיה שנותרו ללא מענה. [אנגלית]
מקור 31 – מדע מפתיע: לא לכל התאים שלנו יש את אותו דנ"א [אנגלית]
מקור 32 – "ביולוגית תא מבוססת ראיות, עם כמה השלכות למחקר הרפואי" דף 29 . [אנגלית]
מקור 33 – רשימה של אורגניזמים לפי מספר הכרומוזומים [אנגלית]
מקור 34 – הלפרן, מקלינטוק "להגדיר את הגנום הבלתי יציב" – Halpern ME. Barbara McClintock on defining the unstable genome. Genetics. 2016; 204:3-4. [אנגלית]
מקור 35 – הבהרת נתיב הביוסינתזה של מאגר חומצות גרעין דנ"א חדש – Genetics: Biosynthesis pathway of a new DNA nucleobase elucidated. [אנגלית]
מקור 36 – מדענים מזהים בסיסים שביעי ושמיני של דנ"א. [אנגלית]
מקור 37 – לסלק את הקללה, הראיות המדעיות לסיום הזיית קוביד – Tom Cowan Breaking the Spell. [אנגלית]
מקור 38 – מה עושה, ולא עושה, אותנו חולים – What Does—and Doesn’t— Make Us Sick. [אנגלית]
מקור 39 – וירולוגיה, הראיות המפלילות – Virology the damning evidence by dpl. [אנגלית]
מקורות כלליים נוספים
בהם נעזרתי לצורך כתיבת מאמר זה:
- הילמן:
- ג'יימי על דנ"א וגנטיקה:
- עומר ג'ורדן על טביעת אצבע בדנ"א:
- הורוס על דנ"א:
- דואון לסטר על גנטיקה: